Trachelipus rathkii Brandt, 1833
 © Агеев Валентин / iNaturalist.org. Минск. CC BY-NC 4.0
© Агеев Валентин / iNaturalist.org. Минск. CC BY-NC 4.0
Семейство Trachelipodidae.
Указывался для Беловежской пущи (Kuznetsova, 2012), а также для "Налибокской пущи". Вероятно, вид распространен значительно шире, так как обнаружен в различных лесах Беларуси. Кроме того, указан и для г. Минска (Островский, 2021).
Является одним из самых широко распространенных видов мокриц России и одним из самых обычных видов мокриц средней полосы Европы и Северной Америки. Наиболее многочислен в лесной и лесостепной зонах.
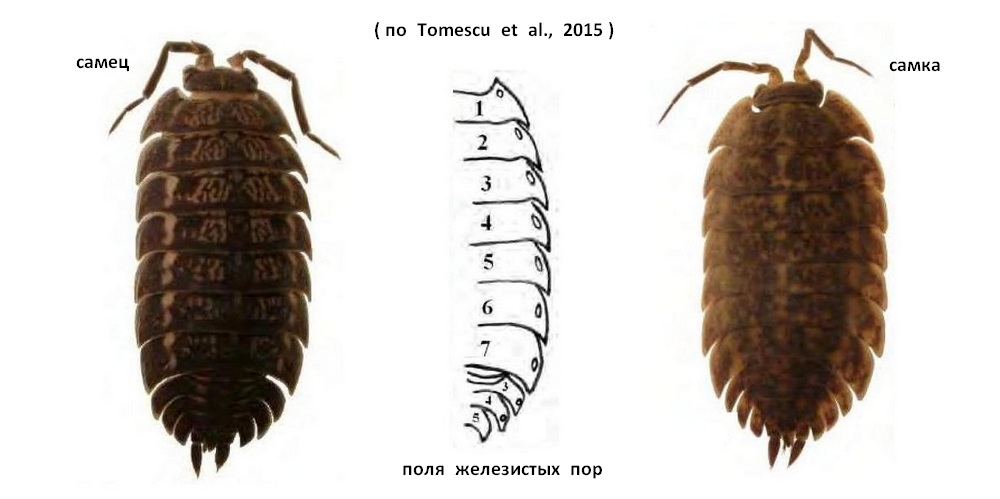
Средняя лопасть головы короткая, выгнутая; боковые лопасти выступают вперед немного дальше. Поля трахей широкие, с бороздками и вздутиями; наружный край трахейного поля зазубрен. Крупные поля пор на всех тергитах расположены посередине края эпимеров. Бугорчатость тергитов выражена слабо. Более подробное описание указано ниже. Окраска изменчива. Чаще встречаются особи с коричневой или сероватой окраской и более светлым рисунком, который соединяется в продольные полосы по бокам тела. Дорсальная часть тела у самцов темно-коричневая. На основании коксальных пластинок имеются желто-оранжевые пятна, которые вместе образуют боковые продольные линии на сегментах переона. На тергитах отчетливый желтоватый рисунок. У самок тергиты светло-коричневые, с желтоватым рисунком. Размеры самцов в Румынии от 6,5х3 мм до 11,5х5 мм; самок: от 7,3х3 мм до 13,5х6 мм. Самые маленькие взрослые особи были собраны в дельте Дуная. Для территории бывшего СССР длина тела указывалась 12-15 мм.
 © Артем Островский, г. Гомель. Все права защищены
© Артем Островский, г. Гомель. Все права защищены
Жгутик антенн состоит 2 крупных члеников. Последний членик антенны длиннее предпоследнего. Боковые головные доли относительно короткие, наклонены латерально. Средняя лопасть головы короткая, выгнутая; боковые лопасти выступают вперед немного дальше. Между боковыми и средней долями имеются косые угловые промежутки. Задние стороны коксальных пластинок тергитов I-III образуют слабую извилистость, коксальные пластины изогнуты непосредственно к задней части тела на IV-VII тергитах. Задние края; эпимеров I тергита переона с глубокой вырезкой. Поля железистых пор овальные на II-VII сегментах и круглые на I сегменте. Они расположены на небольшом расстоянии от боковой стороны коксальных пластинок. Бугорчатость тергитов выражена слабо. На III и IV эпимерах плеона имеются очень маленькие поля железистых пор, очень близко расположенные к внешним сторонам эпимеров. У некоторых самцов они очень трудно заметны, также как у самок, у которых сегменты плеона светло-коричневые с многочисленными желтыми пятнами. Дистальная половина плеотельсона короткая, его боковые стороны образуют косой угол на границе своих проксимально широких и дистально узких половин. Карпоподит VII переопода самца с возвышающимся ребром в проксимальной половине членика имеющим в своей высшей точке округло-тупоугольный выступ, его проксимальная сторона слегка изогнута, а дистальная прямая. Длина гребня не превышает ½ длины карпуса. Ишиоподит булавовидный, с вентральной стороны слегка вогнутый. Переоподы I-VI не имеют специфических морфологических признаков. У переопод I-III имеются плотные ряды шипов на мерусе и карпусе, а на переоподах IV-VI их меньше. На плеоподе I у самца вершина экзоподита короткая и относительно толстая. Экзоподит I плеопода самца с короткой, не доходящей до конца генитальной апофизы; поля пор расположены обычно у края тергитов. Концы эндоподитов имеют ряды коротких шипиков. Плеоподы II-V не имеют специфических морфологических признаков. На экзоподитах плеоподов 5 пар трахей. Поля трахей широкие, с бороздками и вздутиями; наружный край трахейного поля зазубрен. Задний край экзоподита I плеопода самца с узким довольно коротким направленным наружу концом.
Вид экологически пластичен, способен переносить как засуху, так и избыточное увлажнение, а также обладает высокой плодовитостью. В Беларуси выявлен как в хвойных, так и в широколиственных лесах. Найден также в смешанных лесах, но с преобладанием лиственных пород. Самый обычный представитель мокриц юго-востока Беларуси, встречающийся повсеместно, но предпочитающий хорошо дренированные биотопы с сомкнутым травостоем или богатым листовым опадом (уловистость достигает 13,1 экз./м²). Обнаружен в лесных стациях, на лугах, газонах и во дворах застроенной части населенных пунктов. Необходимо отметить, что в застроенной части крупных населенных пунктов мокрица встречается несколько чаще, чем на пойменных лугах и в заболоченных лесах. В то же время на приусадебных участках городов данный вид избегает участков интенсивной и частой обработки почвы и тяготеет к зонам садов с густым травостоем, создающим необходимую для нормальной жизнедеятельности мокрицы повышенную влажность в припочвенном слое воздуха. При этом не обнаруживается предпочтения влажности и механического состава почвы, мокрица многочисленна даже на асфальте у наружных стен зданий под высокотравьем (уловистость составляет 17,6-21,4 экз./м²).
В июле 2019 г. в черте г. Минска за период учетов было собрано и определено 186 экземпляров (7 видов), среди них доля T. rathkii составила 16,13% и вид был четвертым по численности. Это аборигенный для средних широт северного полушария вид, отличающийся выраженной экологической пластичностью. Он обнаружен во всех обследованных биотопах, населяя лесопарковые зоны, газоны и дворы застроенной части Минска. Однако, несмотря на свое широкое распространение, данный вид явно тяготеет к хорошо дренированным биотопам с сомкнутым травостоем или богатым листовым опадом (уловистость достигает 12,8 экз./м²), которые, по большей части, сохранились в частном секторе г. Минска. На приусадебных участках частного сектора г. Минска T. rathkii избегает мест интенсивной и частой обработки почвы, предпочитая сады с густым травостоем, создающим необходимую для нормальной жизнедеятельности мокрицы повышенную влажность в приземистом слое воздуха.
 © semion_levy / iNaturalist.org. Борисовский р-н, Минская обл. CC BY-NC 4.0
© semion_levy / iNaturalist.org. Борисовский р-н, Минская обл. CC BY-NC 4.0
Космополитный эвритопный вид, обитающий в местах с различными экологическими характеристиками. Раду (1985) считает, что это эвритопный вид, обитающий в местах с различной влажностью, под подстилкой, упавшими стволами деревьев, камнями и т. д. Шмидт (1997) утверждает, что Т. rathkii предпочитает открытые места обитания, которых нет в лесах. Томеску и др. (2015) указывают, что собрали представителей этого вида в открытых местах обитания, на лугах с кустарниками или редкими деревьями, в разреженных лесах с песчаной и сухой почвой, в яблоневых и других садах. Экологическое разнообразие мест обитания, в которых был собран этот вид, подтверждает его экологическую пластичность. Для центральной России указывается, что вид способен переносить как засуху, так и избыточное увлажнение, а также обладает высоким коэффициентом размножения. В России наиболее распространен в лесах. В г. Калуга вид обнаружен на всех изученных пробных площадях, но наиболее многочислен он был в биотопах, включающих участки с густым травостоем (луга, влажные насаждения овражно-балочной сети, некоторые сады и дворы). В садах Калуги мокрица избегает обрабатываемых участков.
Характеристика вида как убиквиста является не вполне точной. Несмотря на широкое стациальное распространение, данный вид тяготеет к хорошо дренированным биотопам с сомкнутым травостоем или богатым листовым опадом.
 © Yury Kvach / iNaturalist.org. Лидский р-н, Гродненская обл. CC BY-NC 4.0
© Yury Kvach / iNaturalist.org. Лидский р-н, Гродненская обл. CC BY-NC 4.0
По соотношению полов обследованные биотопы в г. Калуга дифференцируются на две группы. Во всех луговых стациях преобладают самки. В защитных линейных древесных насаждениях преобладают самцы. В биотопах со сложной пространственной структурой соотношение полов в целом недостоверно отклоняется от 50%. Однако здесь наблюдаются представляющие значительный интерес микростациальные различия в соотношении полов. В приусадебном саду самцы преобладают на участках под деревьями и ягодными кустарниками, а самки составляют большинство мокриц в травянистых участках сада. С этой же закономерностью можно связать различия в соотношении полов на садово-огородных участках: в саду 1 с большим обилием кустов черной смородины резко преобладали самцы, в то время как в саду 2 с незначительным участием ягодных кустарников соотношение полов не отличалось от 50%.
Пространственное распределение полов среди мокриц в условиях г. Калуги подчиняется следующей закономерности: на участках с развитой травянистой растительностью преобладают самки, а на участках под деревьями преобладают самцы. При этом обследованные участки городской древесной растительности не обладают ни развитым травостоем, ни лесной подстилкой, которые выполняли бы функцию убежища. Приуроченность самок к участкам с луговой растительностью можно объяснить благоприятными микроклиматическими условиями - высокой относительной влажностью воздуха под травостоем. Для других видов мокриц выявлено, что самки сильнее привлекаются убежищами с высокой влажностью воздуха, чем самцы. Это связывают с необходимостью вынашивания самками яиц. В микростациях с древесной растительностью, но развитым травостоем преобладали самки. Под небольшими группами деревьев среди лугового биотопа наблюдалось такое же соотношение полов, как и на окружающем лугу. Это свидетельствует против избегания самками листового опада деревьев как таковых. Определенное избегание самками участков под смородиной и яблоней в садах объяснить сложнее. Возможно, в этом случае определяющую роль играет низкая температура воздуха и поверхности почвы, а поведение самок направлено на поиск теплых участков, ускоряющих развитие яиц в сумке.
 © halinaa / iNaturalist.org. Жодино, Минская обл. CC BY-NC 4.0
© halinaa / iNaturalist.org. Жодино, Минская обл. CC BY-NC 4.0Во всех стациях г. Калуги обнаружена закономерная сезонная динамика соотношения полов. В мае и июне во всех биотопах, включая луга, преобладают самцы. В июле доля самцов резко снижается, и с начала августа до конца полевого сезона повсеместно преобладают самки. Ранней весной в большинстве биотопов также преобладают самки. Близкий ход имеет сезонная динамика соотношения полов этого и других видов в иных регионах Европы. Так, в Германии доля самцов наиболее велика в мае - начале июня, в то время как осенью, в период низкой плотности преобладают самки. В Великобритании самцы разных видов преобладают в мае и начале июня. Это связывается с повышенной активностью самцов на поверхности почвы в период размножения.
Самки с яйцами встречаются с июня по август, при этом первый пик активности размножающихся самок преимущественно сформирован крупными особями, второй пик - мелкими особями. Установлена зависимость плодовитости мокриц от длины тела. В Калуге у самок T. rathkii в сумке насчитываются от 7 до 106 яиц (в среднем 35,9).
 © Francois Calame / Galerie du Monde des insectes / www.galerie-insecte.org. Suisse. CC BY-NC 4.0
© Francois Calame / Galerie du Monde des insectes / www.galerie-insecte.org. Suisse. CC BY-NC 4.0
 © Francois Calame / Galerie du Monde des insectes / www.galerie-insecte.org. Suisse. CC BY-NC 4.0
© Francois Calame / Galerie du Monde des insectes / www.galerie-insecte.org. Suisse. CC BY-NC 4.0
Литература
- 1. Залесская Н. Т., Рыбалов Л. Б. "Фауна мокриц (Crustacea, Isopoda, Oniscoidea) Москвы и Московской области" / Почвенные беспозвоночные Московской области. Москва, 1982. С. 170-178.
- 2. Kuznetsova D. M., Gongalsky K. B. "Cartographic analysis of woodlice fauna of the former USSR" / Zookeys. 2012; (176): 1-11.
- 3. Tomescu N., Teodor L. A., Ferenti S., Covaciu-Marcov S.-D. "Trachelipus species (Crustacea, Isopoda, Oniscidea) in Romanian fauna: morphology, ecology, and geographic distribution" / North-Western Journal of Zoology 11 (Supplement 1): S1-S106 © NwjZ, Oradea, Romania, 2015.
- 4. Schmidt, C. (1997): Revision of the European species of the genus Trachelipus Budde-Lund, 1908 (Crustacea: Isopoda: Oniscidea). Zoological Journal of Linnean Society 121: 129-244
- 5. Radu, V.G. (1985): Fauna R. S. R.. Crustacea. vol. IV, Fascicola 14 Ordinul Isopoda, Subordinul Oniscoidea, Crinochaeta. Ed. Academiei R. S. R. Bucharest.
- 6. Алексанов В. В. "Особенности биологии мокрицы Trachelipus rathkii (Isopoda, Oniscidea) в условиях города Калуги" / Зоологический журнал. Том 95. №2. 2016. С. 167-180.
- 7. Алексанов В. В. "Соотношение полов в группировках мокрицы Trachelipus rathkei Brandt (Crustacea, Isopoda) в городе Калуге" / Современное состояние и пути развития популяционной биологии: Материалы Х Всероссийского популяционного семинара. Ижевск, 2008. С. 385-387.
- 8. Алексанов В. В. "Распределение мокриц (Isopoda, Oniscoidea) в городе Калуге" / Урбоэкосистемы: проблемы и перспективы развития: материалы III международно научно-практической конференции. Вып. 3. Ишим, 2008. С. 167-169.
- 9. Maksimova S. "5.1.2. Species richness of soil invertebrates" / Naliboki Forest: Land, Wildlife and Human. Volume II. Wild animals. Minsk, 2016. p. 450.
- 10. Максимова С. Л. "Видовой состав мокриц (Crustacea, Isopoda, Oniscoidea) в Беларуси" / Весцi Нац. АН Беларусi. Сер. бiял. навук. 2005 №1. с. 104-106.
- 11. Островский А. М. "К фауне мокриц (Crustacea: Isopoda: Oniscidea) юго-востока Беларуси" / Трансформация экосистем. 2019. Т. 2. № 2 (4). С. 69-78.
- 12. Островский А. М. "Фауна и распределение мокриц (Сrustacea: Isopoda: Oniscidea) в городе Гомеле" / Системы контроля окружающей среды. 2019. Вып. 2 (36). С. 107-116.
- 13. Островский А. М. "Предварительные результаты изучения мокриц (Oniscidea) г. Минска" / Проблемы трансформации естественных ландшафтов в результате антропогенной деятельности и пути их решения: сб. науч. тр. по материалам Междунар. науч. экол. конф., Краснодар, 29–31 марта 2021 г. Краснодар, 2021. С. 311–313.