Gammarus lacustris Sars, 1863
 Photo © CBG Photography Group, Centre for Biodiversity Genomics. v.3 BOLD Systems. CC BY-NC-SA (2009)
Photo © CBG Photography Group, Centre for Biodiversity Genomics. v.3 BOLD Systems. CC BY-NC-SA (2009)
Семейство Gammaridae.
Населяет водоемы и водотоки всех областей Беларуси. Типичный озерный космополит G. lacustris, широко распространенный в Голарктике. В последних исследованиях в нашей стране встречался в местах сбора как в текучих водах, так и в озерах: канал Коссовский, реки Муховец и Ясельда в Брестской обл.; реки Адров и Бирвета, озера Белое и Освейское в Витебской обл.; реки Очесса, Бобрик, Тремля и Неначь в Гомельской обл.; реки Ивенка, Лахва в Могилевской и р. Лучайка в Минской обл.
Длина достигает 1-2,4 см, причем самки меньше самцов. Длина всех особей, отловленных в Беларуси, включая и неполовозрелых 4,5-22,1 мм (9,5±0,3); длина самцов 4,5-22,1 мм (10,1±0,4); длина всех самок 5,4-18,3 мм (8,9±0,3); длина самок без яиц 5,4-18,3 мм (8,9±0,3).
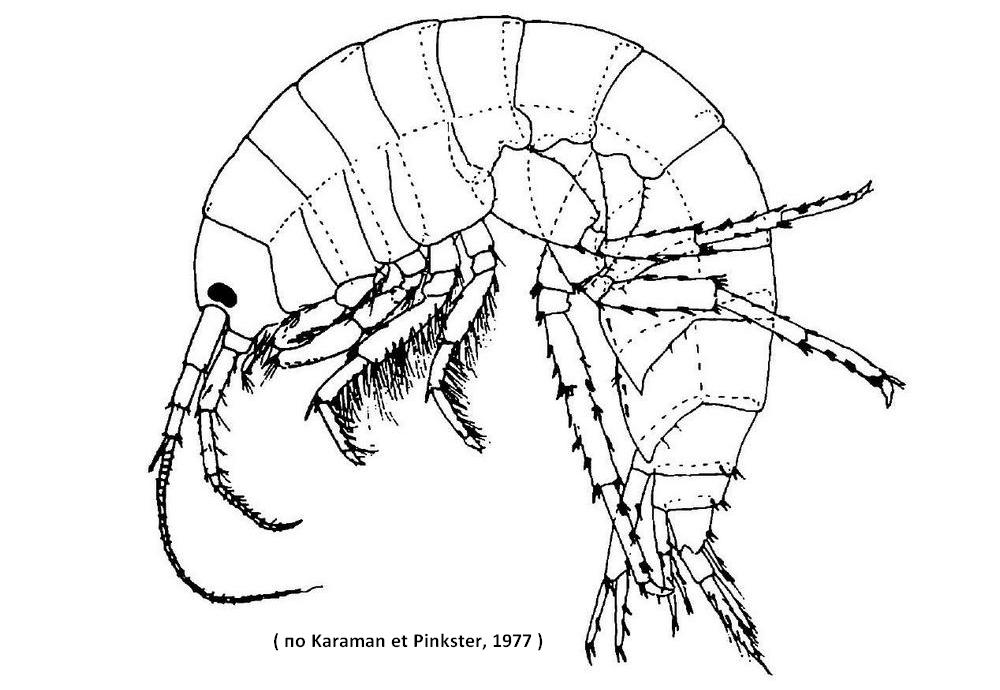
Тело сжато с боков, спинная его сторона дугообразно выгнута. Живые бокоплавы имеют зеленоватую окраску, которая обеспечивается каротиноидами, содержащимися в поедаемых рачками растениях. В целом окраска варьирует от коричневой до сероватой и зависит от среды обитания. Небольшая голова представляет собой синцефалон, в состав которого вошел первый грудной сегмент. По бокам ее передней части расположены фасеточные глаза бобовидной формы. Семь сегментов, входящих в состав грудного отдела свободны. Карапакс отсутствует. По бокам четырех передних грудных сегментов расположены крупные эпимеры - боковые пластинки. Эпимеры остальных сегментов слабо развиты. Брюшной отдел короче грудного и состоит из шести сегментов. Три передних брюшных сегмента имеют одинаковые с грудными размеры и несут плеоподы - плавательные ножки. Эпимеры на этих сегментах выражены слабо. Этот отдел брюшка называется метасома, или плеосома. Задние три сегмента имеют небольшие размеры и лишены эпимеров. Конечности, расположенные на них, отличаются по строению от передних. Этот абдоминальный отдел называется уросома. Брюшко заканчивается тельсоном. Он расщеплен глубокой выемкой на две лопасти, покрытые волосками, и смещен на спинную сторону. Антеннулы двуветвистые. Основная их часть состоит из трех члеников. К ним присоединяется длинная многочленистая дистальная часть. Вторая ветвь короткая и состоит из трех члеников. Антенны одноветвистые, короче антеннул. На антеннулах и антеннах расположены многочисленные чувствительные щетинки и цилиндры. Мандибулы массивные, состоят из базальной части, двух лопастей и трехчленистого щупика. Одна из лопастей - жевательная, сильно хитинизирована и несет по внутреннему краю зубчики. Максиллы I трехлопастные. Наружная лопасть играет роль щупика. Дистальная часть снабжена хитиновыми зубчиками и волосками. Максиллы II состоят из базальной части и двух лопастей с многочисленными щетинками. Конечности первого грудного сегмента, вошедшего в состав сложной головы, превращены в ногочелюсти. Они состоят из шести члеников. На первых двух члениках расположены жевательные лопасти. Первые членики правой и левой пар ногочелюстей срастаются между собой. Семь пар грудных конечностей одноветвистые, их экзоподит редуцирован. В состав каждой конечности входит семь члеников. Коксоподиты прочно срастаются с эпимерами и туловищем. Передние две пары ножек имеют самые маленькие размеры и направлены под острым углом вперед. Их проподит сильно расширен и образует «подклешню», последний членик имеет вид коготка и способен подгибаться к проподиту. Это хватательные конечности - гнатоподы, которые выполняют, главным образом, функцию захвата и удержания пищи. 3-7-я пары грудных конечностей имеют более или менее сходное строение, лишь постепенно по направлению к последнему грудному сегменту увеличиваются в размерах. Коготки третьей и четвертой пар конечностей направлены вперед, последних трех пар - назад. С помощью торакопод бокоплавы способны передвигаться по дну. Кроме того, все кроме первой пары грудных ног несут по одной овальной тонкостенной жаберной пластинке. Они направлены внутрь и прикрыты эпимерами. У самок к коксоподитам 2-5-й пар торакопод прикреплены тонкие хитиновые пластинки - оостегиты, образующие выводковую сумку. Здесь происходит развитие яиц. Из яиц выходят молодые рачки, отличающиеся от взрослых только размерами. Брюшные конечности имеют двуветвистое строение. Передние три пары, расположенные на метасоме (плеосоме), являются плавательными ножками - плеоподами. Они состоят из протоподита и причленяющихся к нему многочленистых экзо- и эндоподитов, покрытых многочисленными волосками. С помощью плеопод бокоплавы плавают спинной стороной кверху там, где глубина достаточная, а на мелководье - на боку. Благодаря такой особенности бокоплавы и получили свое название. Помимо локомоции плеоподы, постоянно двигаясь, обеспечивают циркуляцию воды, омывающей жаберные пластинки. 4-5-я пары брюшных конечностей, принадлежащих сегментам уросомы, направлены назад и закреплены на теле неподвижно. Они также двуветвистые, но их экзо- и эндоподиты нечленистые и снабжены короткими шипиками. С помощью этих ножек, называемых уроподами, бокоплавы способны прыгать, отталкиваясь от субстрата. Последняя пара уропод более подвижна, чем две передние. Они также имеют двуветвистое строение, но имеют расчлененные экзо- и эндоподиты, покрытые щетинками.
 Photo by User:Vikte at Wikimedia Commons. CC BY-SA 4.0
Photo by User:Vikte at Wikimedia Commons. CC BY-SA 4.0
G. lacustris обитает в различных озерах Северного полушария, иногда достигая высокой численности. Обычно считается обитателем стоячих вод, хотя в Польше локально встречается в некоторых довольно быстрых ручьях и реках, которые не связаны с озерами. Сообщается о подобных находках и в шведской Лапландии. Однако озера, водохранилища и пруды являются основной средой обитания.
Данные о кислородном режиме, отношению к щелочности и солености воды также противоречивы. Если ранее указывалось, что из-за чувствительности к содержанию в воде кислорода не живет в заиленных и загрязненных водоемах и по этой же причине содержание в аквариумах представляет определенные затруднения; то несколько позже, что вид толерантен к низким концентрациям кислорода и высоким концентрациям ионов. В центральной части России G. lacustris обитает в озерах, в воде которых содержится 9 г сухих остатков на литр. В центральной Польше (Лодзинское воеводство) этот вид обитает в солоноватых торфяных болотах, где содержание хлоридов составляет более 3 г на литр. Может длительное временя жить при низкой температуре, но довольно чувствителен к температурам выше 20°C.
 Photo © CBG Photography Group, Centre for Biodiversity Genomics. v.3 BOLD Systems. CC BY-NC-SA (2009)
Photo © CBG Photography Group, Centre for Biodiversity Genomics. v.3 BOLD Systems. CC BY-NC-SA (2009)
Жизненный цикл точно не известен. В Великобритании новое поколение, появившееся в текущем году, начинает размножаться зимой, а молодое поколение появляется в апреле и мае. В Польше весеннее поколение, по-видимому, не размножается в конце лета или осенью того же года. Возможно, период размножения этого вида в Польше несколько смещен во времени из-за более суровых зим, но точные наблюдения отсутствуют. Можно также отметить, что размножающиеся самки в Норвегии отмечаются до сентября. Для Великобритании указывается годичный жизненный цикл вида; для канадских ручьев (подвид limnaeus) описывается двухлетний жизненный цикл, в котором наиболее интенсивное размножение происходит в мае, июне и июле. Эта разница в жизненном цикле может быть связана со значительно более низкими температурами этих водных потоков в течение года (всегда ниже 13°C). В целом, для средней Европы приняты следующие данные для вида: средний размер размножающихся самок 11,28 мм, средний размер кладки 18,75 яиц, период размножения 4 месяца, 1 генерация в год.
Питается растениями, как живыми, так и отмершими, могут использовать в пищу также останки животных.
 Photo © CBG Photography Group, Centre for Biodiversity Genomics. v.3 BOLD Systems. CC BY-NC-SA (2011)
Photo © CBG Photography Group, Centre for Biodiversity Genomics. v.3 BOLD Systems. CC BY-NC-SA (2011)
Литература
1. Круглова О. Ю. "Класс Crustacea – Ракообразные : метод. рекомендации". В 2 ч. Ч. 2. Высшие ракообразные. Минск, 2014. -43с.
2. Макаренко А. И. "Размерные характеристики чужеродных и аборигенных видов амфипод в Беларуси" / Весці Нацыянальнай Акадэмii Навук Беларусі. Серыя біялагічных навук. № 1, 2015. С.100-105
3. Jażdżewski Krzysztof "Remarks on Gammarus lacustris G. O. Sars, 1863, with description of Gammarus varsoviensis n. sp. (Crustacea, Amphipoda)" / Bijdragen tot de Dierkunde, 45 (1). 1975. pp.71-86
4. Rewicz T., Grabowski M., MacNeil C., Bącela-Spychalska K. "The profile of a ‘perfect’ invader – the case of killer shrimp, Dikerogammarus villosus" / Aquatic Invasions (2014) Volume 9, Issue 3: 267–288
5. Макаренко А. И., Вежновец В. В. "Современное распределение чужеродных и аборигенных видов отряда Amphipoda Latreille, 1816 на территории Беларуси" / Весцi Нацыянальнай Акадэмii навук Беларусi. Серыя бiялагiчных навук. № 4, 2014. С.95-99
6. Karaman G.S. & Pinkster S. (1977). Freshwater Gammarus species from Europe, North Africa and adjacent regions of Asia (Crustacea - Amphipoda). Part I. Gammarus pulex - group and related species. Bijdragen tot de Dierkunde. 47, 1, 1-97.