Eurytemora velox Lilljeborg, 1853
 Photo András Gubányi, Katalin Kovács, László Forró / Hungarian Natural History Museum. CC BY-NC-ND 3.0
Photo András Gubányi, Katalin Kovács, László Forró / Hungarian Natural History Museum. CC BY-NC-ND 3.0
Отряд Calanoida. Семейство Temoridae.
Впервые (1956) в водоемах нашей республики эуритемора велокс обнаружена в р. Припять и ее притоках на участке от г. Пинска до устья. Позже ее обнаружили и в реках Пина, Сколодинка и затоке в пойме Припяти, выше впадения р. Уборть. В 1970 г. зафиксировано довольно широкое распространение этого вида в придаточных системах и устьевых участках притоков р. Припять: ниже Найдобелянского канала, выше р. Уборть, ниже р. Сколодинки, ниже р. Брагинки и в устье самой Припяти. В 1996 г. подтверждено нахождение вида в р. Пина 12,2 км выше Пинска, в 1997 - в Днепро-Бугском канале у п. Дубои. С 1983 г. установлено распространение эуритеморы в реках бассейна среднего Днепра, а в 1996–97 гг. - в р. Сож, вблизи Гомеля. В это же время рачок найден и в р. Мухавец, выше Кобрина. По сборам за период с 1996 по 2011 г. подтверждено нахождение E. velox в реках Западный Буг, Припять и Сож и их придаточных водоемах.
Самка. Тело более или менее короткое, коренастое, с наибольшей шириной на границе цефалона и 1-го сегмента торакса. Последний торакальный сегмент с большими, направленными назад крыловидными выростами, внешние края выростов волнисто изогнуты и вооружены тонкими шипиками. Абдомен симметричный, укороченный, его длина равна приблизительно 2/3 длины цефалоторакса. На анальном сегменте шипиков нет. Каудальные ветви короткие, они значительно короче двух последних абдоминальных сегментов, вместе взятых; внутренние края каудальных ветвей опушены волосками, иногда на дорсальной поверхности видны мелкие шипики. Каудальные щетинки короткие и густо оперены волосками. Антеннулы короткие; они едва достигают последнего торакального сегмента или даже еще короче. Строение околоротовых частей и 1-4-й пар плавательных ног обычное для видов рода. Базиподит 5-й пары ног короткий и широкий, его ширина в 1,5 раза больше длины и несет короткую щетинку. 1-й членик экзоподита примерно в полтора раза длиннее базиподита, с относительно коротким заостренным внутренним выростом, направленным назад под углом 45° и вооруженным дистально несколькими мелкими шипиками; на середине наружного края 1-й членик экзоподита с 1 голым шипом. 2-й членик экзоподита маленький, овальный и вооружен 2 шипами: латеральный шип голый, в 2,5 раза короче апикального, апикальный шип густо покрыт мелкими шипиками в дистальной половине. Длина самок 1,3-2,2 мм.
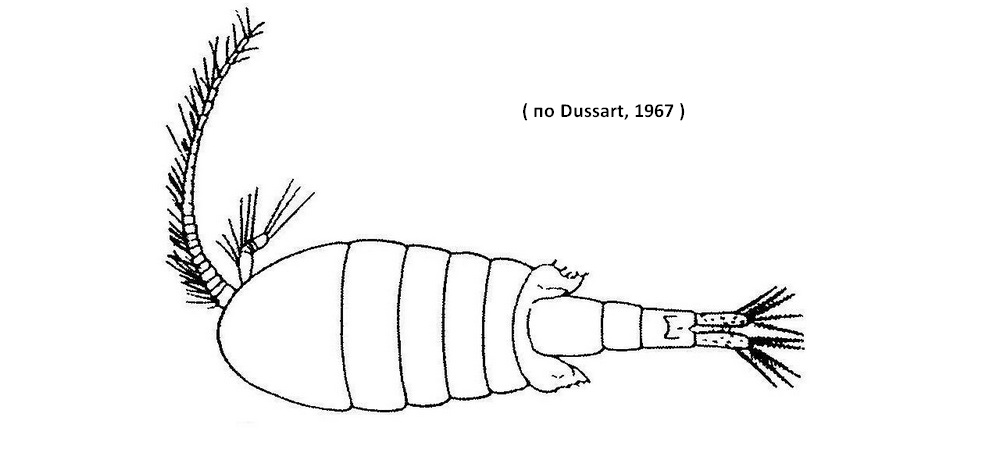
Самец. Тело значительно более тонкое и стройное, чем у самки; крыловидных выростов на последнем торакальном сегменте нет. Каудальные ветви и каудальные щетинки длиннее по сравнению с таковыми у самки - длина каудальных ветвей приблизительно равна общей длине двух последних абдоминальных сегментов. Правая антеннула геникулирующая, 21-члениковая. 8, 9 и 12-й членики несут по загнутому шипу, из которых шип на 8-м членике самый маленький; 10-й и 11-й членики ничем не вооружены. Правая нога 5-й пары 5-члениковая; базиподит этой ноги удлинен, внутренний край его заметно выпуклый, внешний почти прямой; 1-й членик экзоподита длинный,в дистальной части вооружен парой мелких шипиков; 2-й и 3-й членики экзоподита иногда отделены друг от друга не вполне ясно, но все же установить между ними границу возможно всегда; 2-й членик экзоподита несет 2 щетинки в дистальной части; 3-й членик вооружен 1 или 2 щетинками по внутреннему краю. Базиподит левой ноги 5-й пары широкий и короткий, с большим округлым выступом по внутреннему краю; 1-й членик экзоподита примерно в полтора раза длиннее предыдущего, несет маленький терминальный шип; 2-й изогнут под тупым углом и лопатообразно расширен на конце, образуя двураздельную лопасть, вооруженную очень мелкими шипиками и волосками; внешний и внутренний края 2-го членика экзоподита несут по 1 шипу. Длина самцов 1,2-1,8 мм.
Тело более или менее прозрачное, с желтовато-коричневым оттенком.
Форма эвригалинная, диапазон выносимой солености очень велик. В континентальных водах вид встречается в различных водоемах: от временных до озер с повышенной соленостью, переживая неблагоприятные условия на всех стадиях развития, включая стадию яйца. Основные места обитания сосредоточены вблизи устьев рек, где он создает достаточно большие плотности и является кормовым объектом для карповых рыб. Кроме того, встречается в различных пресных водоемах Европы - от Норвегии до устья Дуная, от Англии на западе, до Урала - на востоке. Главнейшие места обитания - солоноватые воды, однако, как указано выше, вид активно осваивает пресные воды Беларуси, хотя и встречается в небольшом количестве в водоемах Полесья от стариц до основного русла рек.
Вид относительно недавно проник в водоемы юга республики и его ареал постепенно расширяется. Этому процессу способствует эврибионтность и приспособленность вида к переживанию неблагоприятных условий (высыхание, промерзание) даже на поздних стадиях развития. Нестабильное гидрологическое состояние таких экосистем, как пойменные водоемы и реки, особенно в условиях зарегулирования, способствует обширному расселению чужеродных видов, в том числе и E. velox. Успеху этого процесса также способствует предпочтение E. velox прибрежных зон водоемов и водотоков, то есть участков с замедленным течением, в условиях которых возможно самостоятельное передвижение против течения. Благодаря высокой степени адаптации к изменениям среды обитания, не исключается расселение и пассивным способом - переносом другими животными.
В стоячих водоемах при обследовании разных местообитаний для этого вида обнаружено предпочтение прибрежных биотопов, где средняя численность рачка была в 10 раз выше, чем в пелагической части водоема. Эта особенность распределения позволяет эуритеморе занять отличную от других каланоидных копепод пространственную нишу. При этом максимальная плотность создается в открытом прибрежье. Кроме этого, наблюдаются возрастные различия в заселении. В чистой (незаросшей) литорали преобладают рачки 1-4-й стадий развития, а заросли предпочитают более взрослые особи. E. velox в текучих водах приурочена к заросшему (защищенному) прибрежью, где ее плотность в 6 раз больше, чем в открытых участках. В пойменных озерах анализ биотопической приуроченности этого вида показал, что основная масса популяции также держится в прибрежье, а численность на порядок превышает таковую на пелагических станциях. Анализ возрастного состава вида в двух основных типах литоральной зоны (свободной от высшей водной растительности и заросшей тростником) показал четкие различия в распределении рачков разных возрастов в этих двух биотопах. В чистой (не заросшей) литорали преобладали рачки младших копеподитных стадий (I-IV), заросли предпочитали более взрослые особи (V и VI копеподитных стадий).
Cредняя абсолютная плотность эуритеморы для двух обследованных станций на р. Мухавец - 785 экз./м³, средняя относительная плотность (от общей численности зоопланктона) - 0,227 %. Количество эуритеморы от общей численности копепод было максимальным в заросшем прибрежье Мухавца в д. Бульково - 360 экз./м³ (0,34 %), минимальным - в чистом прибрежье Мухавца в Бресте (40 экз./м³, или 0,013 %). В целом, плотность рачка в Мухавце у д. Бульково намного выше, чем плотность его в реке в черте города. Количество рачка на науплиальных стадиях в целом в несколько раз выше, чем взрослых особей. Наиболее высока их плотность в чистом прибрежье реки у д. Бульково - 0,575 % (620 экз./м³). Минимальное - там же в зарослевой зоне (0,284 %, или 300 экз./м³).
В условиях водоемов Беларуси E. velox относится к фильтраторам и специализируется на потреблении диатомовых водорослей прибрежья, предположительно с поверхности обрастаний и дна. Известно, что в зарослевом фитопланктоне озер Беларуси доля диатомей составляет около 55%, а в перифитоне - 85%, и в нем часто встречаются подвижные перифитонные формы, способные переходить к планктонному образу жизни. Высокое флористическое разнообразие диатомовых в эпифитоне водоемов может являться обильным и доступным кормом для копепод. В данном случае питание E. velox диатомеями в литорали помогло, очевидно, найти свою нишу в пресных водах и успешно избежать конкуренции с аборигенными растительноядными видами каланоидных копепод рода Eudiaptomus, обитающими в пелагической части подобных водоемов.
 Photo © A. Hobæk, Norwegian Institute for Water Research. v.3 BOLD Systems. CC BY-NC-SA (2015)
Photo © A. Hobæk, Norwegian Institute for Water Research. v.3 BOLD Systems. CC BY-NC-SA (2015)
Литература
1. Вежновец В. В., Литвинова А. Г., Макаренко А. И. "Особенности мониторинга чужеродных видов ракообразных" / Мониторинг окружающей среды : сб. материалов II международной науч.-практ. конф.: в 2 ч. Ч.2. Брест, 2013. С.22-24
2. Литвинова А. Г. "Количественное развитие чужеродного планктонного вида в реке Мухавец" / Мониторинг окружающей среды : сб. материалов II международной науч.-практ. конф.: в 2 ч. Ч.2. Брест, 2013. С.60-62
3. Боруцкий Е. В., Степанова Л. А., Кос М. С. "Определитель Calanoida пресных вод СССР". Ленинград, 1991. -504с.
4. Вежновец В. В. "Ракообразные (Cladocera, Copepoda) в водных экосистемах Беларуси: каталог. Определительные таблицы". Минск, 2005. -150с.
5. Вежновец В. В., Зайдыков И. Ю., Наумова Е. Ю., Сысова Е. А. "Особенности биологии двух видов копепод (Crustacea, Copepoda, Calanoida) как возможные причины изменения их ареалов" / Российский Журнал Биологических Инвазий. № 2. 2012 С.16-29