Polyphemus pediculus Linnaeus, 1758
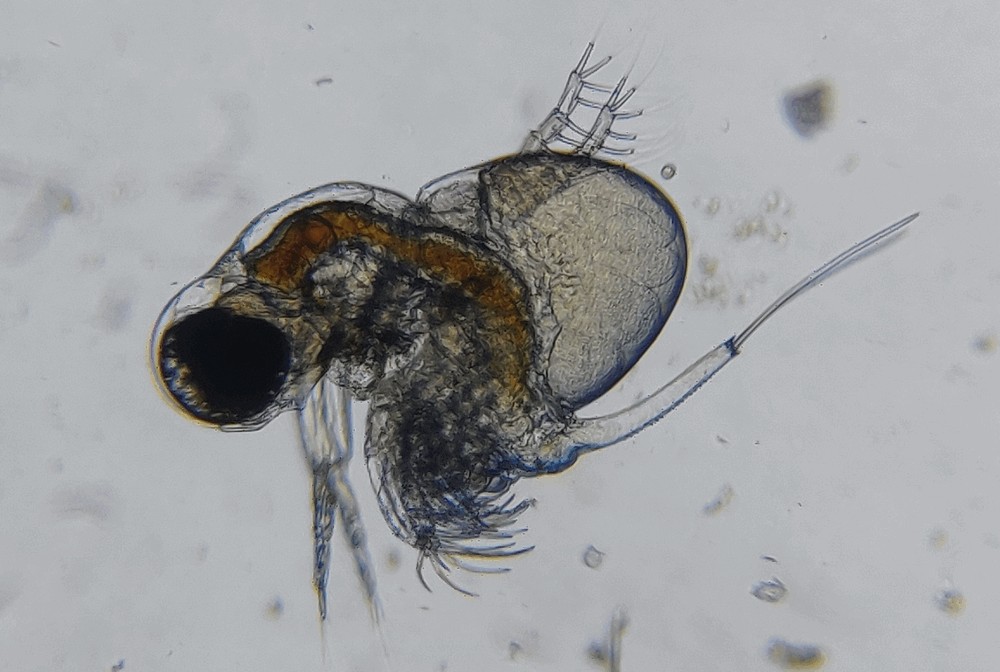 Photo © Nazarov Lev / iNaturalist.org. CC BY-NC 4.0
Photo © Nazarov Lev / iNaturalist.org. CC BY-NC 4.0
Onychopoda. Семейство Polyphemidae.
В водоемах Беларуси встречается повсеместно. Это относится как к озерам и прудам, так и к рекам и водохранищам, как к болотам, так и ко временным водоемам.
Тело прозрачное, серовато-желтой окраски. Раковинка направлена назад, она рудиментарная - в виде мешкообразного выроста, служащего выводковой камерой. Голова подвижная, большая, удлиненная, спереди почти шарообразная, без латеральных килей, отделена от туловища глубоким вдавлением. Тело толстое, короткое. Постабдомен очень короткий, переходящий в довольно длинный хвостовой стебелек, несущий на конце хвостовые щетинки. Передние антенны короткие, почти достигают переднего края головы; их частично слившееся основание прижато к нижнему краю головы. Задние антенны короткие, с 7 плавательными щетинками на каждой ветви. Верхняя ветвь 4-члениковая, нижняя - 3-члениковая. Верхняя губа большая, шлемовидная. Жвалы с крупными зубцами. Четыре пары ног, все пары ног снабжены хорошо развитыми экзоподитами. Абдоминальные коготки отсутствуют. Самцы меньше самок. Последний членик I пары ног снабжен крючком. Семепроводы открываются близ анального отверстия. Длина самки 1,2-1,8, самца 0,9-0,95 мм.
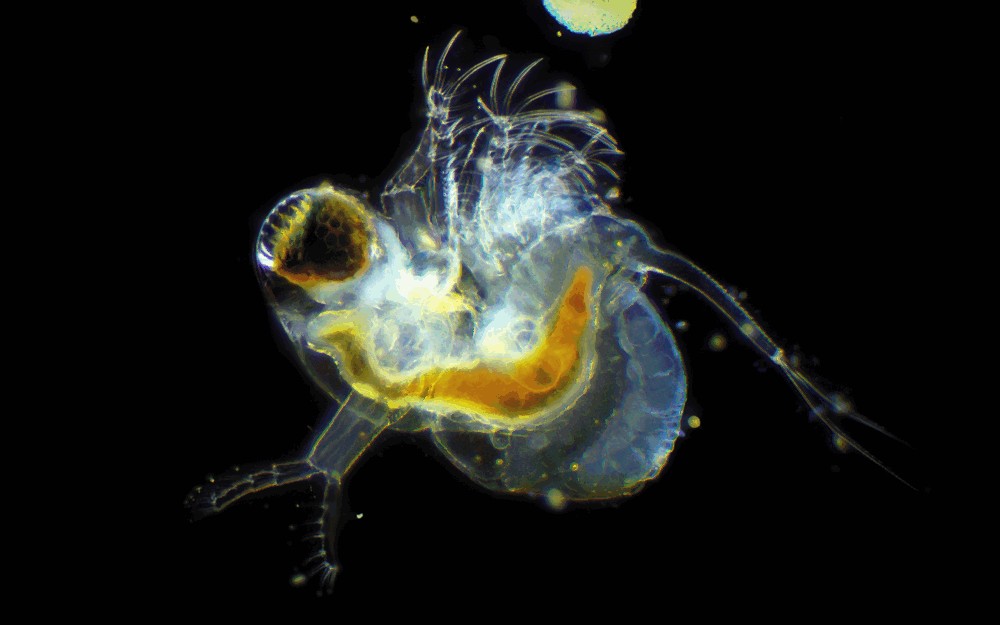 Photo © Ivan / iNaturalist.org. CC BY 4.0
Photo © Ivan / iNaturalist.org. CC BY 4.0
Самка. Дорсальную часть компактного тела занимает раковина, всю внутреннюю полость которой заполняет выводковая сумка, служащая для вынашивания зародышей. Рост раковины происходит в соответствии с ростом зародышей, поэтому у молодых самок раковина мала, а по мере созревания зародышей становится почти шарообразной. Голова продолговатая. Всю ее переднюю часть занимает крупный сложный глаз с большим скоплением пигмента в центре. Дорсальная часть головы занята головным щитом, служащим для дыхания: его поверхность покрыта слоем хитина, под которым хорошо видны шестигранные клетки эподермы. За головным щитом располагается глубокая выемка, отделяющая голову от туловища. Антеннулы прижаты к передне-нижнему краю головы. Они имеют общее основание, а дистально оканчиваются каждая пятью эстетасками. Плавательные антенны сильные и относительно длинные. Они имеют складчатое основание, почти равное по длине первому членику. Ветви антенн: верхняя - четырехчлениковая и нижняя - трехчлениковая. Основной членик антенны со стороны трехчлениковой ветви имеет хорошо заметный вырост. Каждая ветвь несет по 7 оперенных щетинок. Щетинки двучлениковые; нижняя часть их цилиндрическая, верхняя - плоская; щетинки густо оперены по всей длине. Вдоль края ветвей антенн имеется по несколько насечек на каждом членике. Губа с выпуклым передним краем. Мандибулы имеют многочисленные зубцы, сгруппированные в три ряда, среди которых один э два зубца наиболее сильно выдаются. Максиллулы в виде двух маленьких опушенных лопастей. Грудных конечностей четыре пары. Пара I длиннее, чем II и III. Конечности двуветвисты. Экзоподиты плоские и широкие и снабжены на I-III паре пятью оперенными щетинками. Эндоподиты трехчлениковые. Первые членики несут по нижнему краю по два ряда многочисленных оперенных щетинок, число которых в ряду от I пары до III колеблется соответственно: 6-10, 4-7 и 4-5. Вторые членики эндоподитов несут по две загнутые когтевидные щетинки, а конечные - по 4 когтевидных, слабо оперенных щетинки. IV пара ног небольшая, но имеет хорошо заметный экзоподит с тремя маленькими загнутыми щетинками. Максиллярные придатки различимы на всех четырех парах ног. На I паре - это бугорок с оперенной щетинкой, на II и III - опушенный отросток с небольшими зубцами и на IV паре - маленький выступ с двумя зубчиками. Кауда в виде закругленного бугорка, на вершине которого между двумя анальными лопастями открывается анальное отверстие. Кзади кауда несет длинный цилиндрический вырост - „хвостовой стебель", на конце его располагаются плавательные щетинки. В отличие от других Cladocera они не опушены. Плавательные щетинки обычно заходят за выводковую сумку и выдаются с каудальной стороны тела.
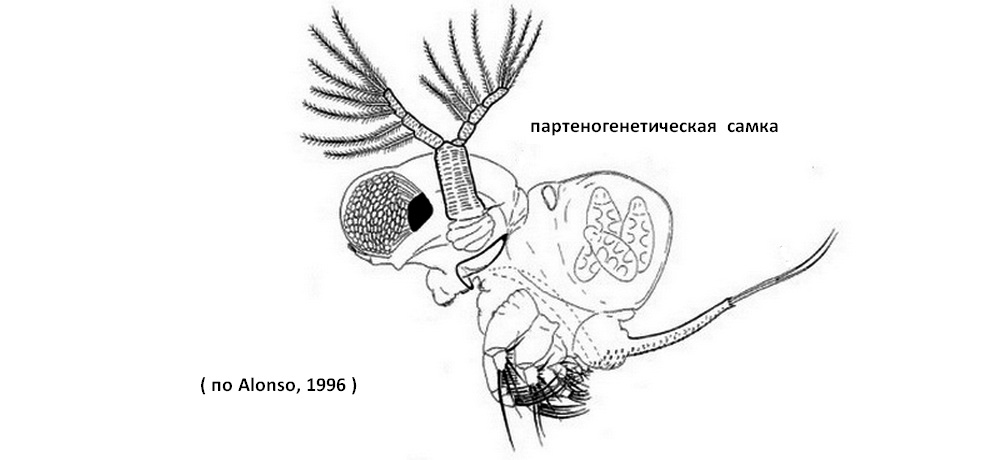 Самки делятся на партено- и гамогенетических. Последние значительно крупнее первых.Партеногенетическая самка имеет слабую серовато-коричневую окраску. Высота самки 1,2-1,8 мм, длина до 0.9 мм. Гамогенетическая самка несет в выводковой сумке зимние яйца и не приспособлена к вынашиванию партеногенетических зародышей. Зимние яйца одеты плотной оболочкой, их количество колеблется от 1 до 9. Наиболее ярко окрашены гамогенетические самки, у которых туловище и ноги имеют красноватые, фиолетовые и синие оттенки, а выводковая камера коричнево-красного цвета. Длина гамогенетической самки до 0,85 мм.
Самки делятся на партено- и гамогенетических. Последние значительно крупнее первых.Партеногенетическая самка имеет слабую серовато-коричневую окраску. Высота самки 1,2-1,8 мм, длина до 0.9 мм. Гамогенетическая самка несет в выводковой сумке зимние яйца и не приспособлена к вынашиванию партеногенетических зародышей. Зимние яйца одеты плотной оболочкой, их количество колеблется от 1 до 9. Наиболее ярко окрашены гамогенетические самки, у которых туловище и ноги имеют красноватые, фиолетовые и синие оттенки, а выводковая камера коричнево-красного цвета. Длина гамогенетической самки до 0,85 мм.
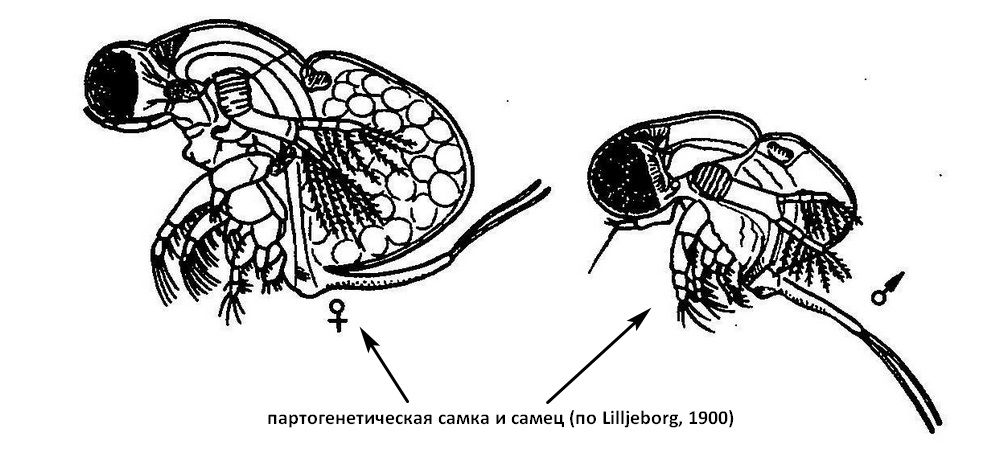 Самец. Раковина редуцирована до небольшого бугорка. Голова занимает больше половины высоты тела. Антеннулы крупные; из пяти эстетасков один вытянут, отогнут в виде щупика („бич"). Третий членик ног I пары значительно видоизменен. Из четырех конечных щетинок у самца имеется только две - короткая и длинная, которые сдвинуты к вентральной стороне членика. Конечная часть членика расширена и уплотнена, образуя „пятку". От верхнего края пятки наискось, острием ко второму членику располагается хватательный крючок с зазубренным внутренним краем. Апикальная часть чпеника имеет небольшую саблевидную щетинку, опушенные бугорки и еще один маленький крючок, также обращенный острием ко второму чпенику. Второй и третий членики эндоподита имеют краевые зубцы, обращенные внутрь. Пенис самца представляет собой небольшое овальное образование впереди анального отверстия, прикрытое IV парой ног. Спереди пенис покрыт волосками, каудальная часть его гладкая. Семепроводы открываются на вершине пениса. Хвостовой стебель самца выдается далеко назад, так как тело его сильно укорочено. Длина половозрелого самца 0,5-0,77 мм.
Самец. Раковина редуцирована до небольшого бугорка. Голова занимает больше половины высоты тела. Антеннулы крупные; из пяти эстетасков один вытянут, отогнут в виде щупика („бич"). Третий членик ног I пары значительно видоизменен. Из четырех конечных щетинок у самца имеется только две - короткая и длинная, которые сдвинуты к вентральной стороне членика. Конечная часть членика расширена и уплотнена, образуя „пятку". От верхнего края пятки наискось, острием ко второму членику располагается хватательный крючок с зазубренным внутренним краем. Апикальная часть чпеника имеет небольшую саблевидную щетинку, опушенные бугорки и еще один маленький крючок, также обращенный острием ко второму чпенику. Второй и третий членики эндоподита имеют краевые зубцы, обращенные внутрь. Пенис самца представляет собой небольшое овальное образование впереди анального отверстия, прикрытое IV парой ног. Спереди пенис покрыт волосками, каудальная часть его гладкая. Семепроводы открываются на вершине пениса. Хвостовой стебель самца выдается далеко назад, так как тело его сильно укорочено. Длина половозрелого самца 0,5-0,77 мм.
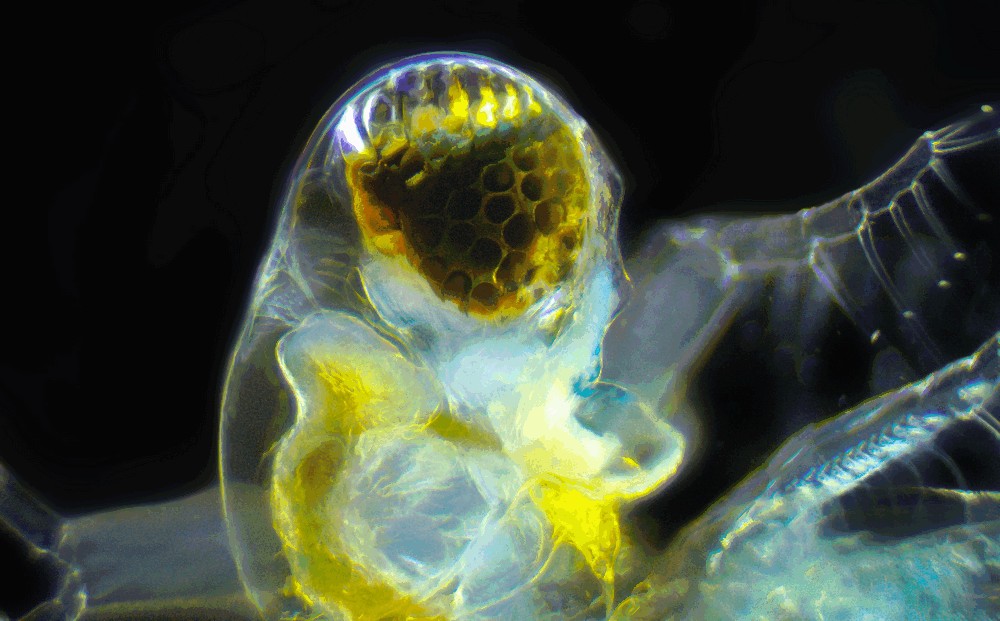 Photo © Ivan / iNaturalist.org. CC BY 4.0
Photo © Ivan / iNaturalist.org. CC BY 4.0
Органы дыхания устроены иначе, чем у прочих ветвистоусых. Конечности хищных форм лишены эпиподитов - дыхательных отростков или жабр. Газообмен совершается исключительно с помощью затылочного органа (так называемого головного щита), представляющего собой скопление крупных железистых клеток. Он в виде большого вздутия располагается за головой выше прикрепления плавательных антенн и примыкает со спинной стороны к сердцу. Здесь наблюдается наибольшая проницаемость покровов, происходят поглощение кислорода и выделение углекислоты. Способ дыхания посредством головного щита - единственный.
Кровеносная система устроена примитивно и состоит из сердца, имеющего пару остий (отверстий), через которые протекает гемолимфа, циркулирующая затем по телу среди мембран и перекладин внутреннего скелета. Сердце представляет собой мешочек, расположенный позади затылочного органа, непосредственно над выводковой сумкой.
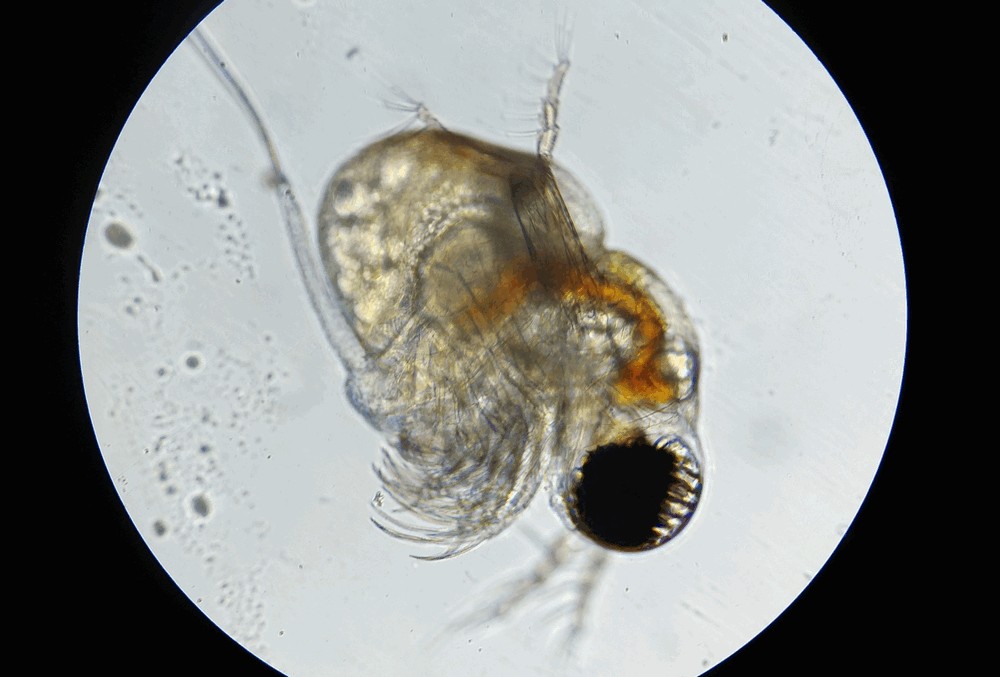 Photo © Nazarov Lev / iNaturalist.org. CC BY-NC 4.0
Photo © Nazarov Lev / iNaturalist.org. CC BY-NC 4.0
Планктонные животные, обладающие ярко выраженными приспособлениями к парению в толще воды. Молодь сразу после рождения ведет планктонный образ жизни. Лишь одна стадия жизненного цикла - латентное (покоящееся) яйцо опускается на дно либо в глубинные слои и находится там до выхода молодого рачка.
Типичный обитатель малых, в том числе временных водоемов и прибрежной зоны крупных. Встречается в водоемах самого различного типа - прудах, временных водоемах, прибрежье рек, озер, водохранилищ, Балтийского и Аральского морей. Предпочитает защищенные места в прибрежье, где может создавать значительные плотности. Полифем предпочитает защищенное прибрежье - свободные окна среди зарослей водной растительности. Область его распространения в озерах и водохранилищах ограничивается глубиной 1,5-2 м, хотя максимальная численность наблюдается у уреза воды от 10 до 40 см. P. pediculus, обитающий на самом мелководье, может держаться ближе или дальше от поверхности воды; на участках с глубиной 1-2 м перемещается по вертикали в течение суток. Отдельные особи выносятся на глубину ветровым течением. Большая часть площади обитания в водоемах входит во временно обсыхающую зону, поэтому латентные яйца переносят промерзание и высыхание. Это делает возможным расселение вида во временные водоемы. Известны случаи распространения далеко от берегов.
Летний, теплолюбивый вид. Появляется в водоеме при температуре 10,5-12°С, и колебания от 11 до 29° не сказываются на интенсивности его партеногенетического размножения. Исчезает он обычно в конце августа, отдельные экземпляры доживают до сентября - начала октября при температуре воды около 11°С. Обычно исчезновение рачка происходит значительио раньше - в июле - начале августа, не вследствие понижения температуры, а после обсыхания мелководных биотопов.
Отношение к общей жесткости воды неясно. Он встречается в самых различных водоемах, в том числе и в кислых, болотистых. Описаны случаи нахождения Р. pediculus в Финском заливе, при солености 12-13‰ и в прибрежных водах Арала, при солености 1-6‰, куда он заносится течением из дельты Сыр-Дарьи.
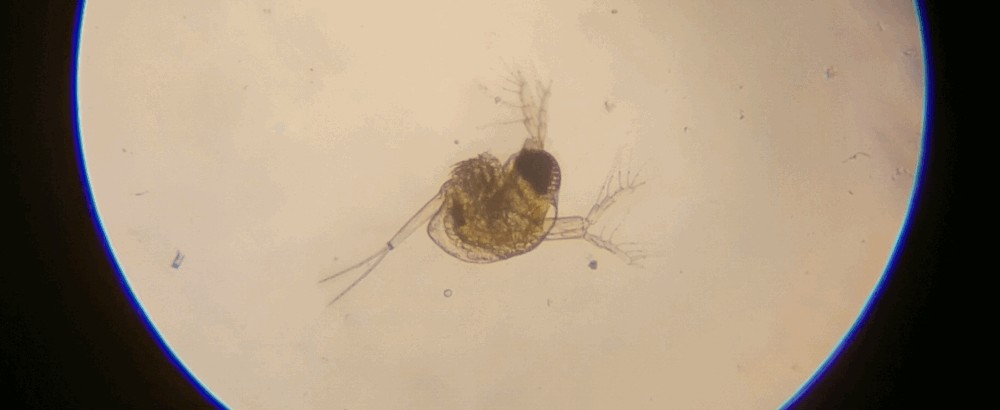 Photo © Mikko Heikkinen / iNaturalist.org. CC BY 4.0
Photo © Mikko Heikkinen / iNaturalist.org. CC BY 4.0
Вид северного происхождения, послеледниковый иммигрант, имеет очень короткий жизненный цикл, приспособленный к арктическому лету. В Рыбинском водохранилище P. pediculus появляется в планктоне в конце апреля и при благоприятных условиях интенсивно размножается, достигая в июне максимальной численности. Затем в июле и августе образуется несколько пиков, однако они ниже первого весеннего. Самцы и гамогенетические самки появляются в популяциях несколько раз, что характерно для этого полицикличного вида. Во второй половине лета численность рачка значительно понижается и к концу августа - сентябрю полифем исчезает, отложив латентные яйца. Иногда популяция погибает раньше - в июле - начале августа, в связи с обсыханием биотопов.
P. pediculus, населяющий мелководья почти от уреза воды, естественно, перемещается по вертикали в небольшом диапазоне. На участках с глубиной 50 см в светлое время суток (от 5 до 19 ч) основная масса рачков концентрируется у поверхности воды, образуя плотные, иногда грибовидные стаи, пятнами разбросанные по всему мелководью. С заходом солнца эти скопления разреживаются, рассеиваются, ночью рачки распределяются по всей толще воды равномерно. С восходом солнца стаи образуются вновь на том же месте. При максимальной освещенности плотность рачков у поверхности уменьшается. Особенно избегают прямого света новорожденные рачки, они мигрируют в нижние, менее освещенные горизонты.
Миграции партеногенетических и гамогенетических самок, а также самцов несколько различаются. Партеногенетические самки так же, как новорожденные рачки, в полдень отходят от поверхности. С наступлением темноты стаи рассеиваются, а перед восходом, с 4 до 6 ч, концентрируются в верхнем 5-сантиметровом слое, где происходит отрождение молоди. В начале лета гамогенетические самки и самцы встречаются единичными экземплярами, и их миграции трудно проследить. В августе особи обоих полов при максимальной освещенности быстро мигрируют от поверхности, В отличие от партеногенетических самок, образующих плотные стаи днем, гамогенетические самки и самцы концентрируются в стаи лишь на рассвете. В остальное время они рассредоточены по всей толще воды.
 Photo © Nazarov Lev / iNaturalist.org. CC BY-NC 4.0
Photo © Nazarov Lev / iNaturalist.org. CC BY-NC 4.0
Обладает, как все Cladocera, двумя способами размножения: партеногенетическим и гамогенетическим, с участием самцов и гамогенетических самок. Партеногенетическое размножение продолжается в течение всего жизненного цикла вида, до исчезновения последнего из водоема и является способом интенсивного наращивания численности популяцией. Гамогенез прослеживается в конце цикла; в результате: двуполого размножения появляются латентные яйца, способные переносить неблагоприятные условия.
Как и у остальных ветвистоусых, новорожденный рачок появляется на свет крупным, полностью развитым. Развитие яйца происходит в замкнутой выводковой сумке, стенки которой постепенно выделяют питательные вещества, необходимые для полного развития зародыша. Выводковая сумка ограничена спереди поперечной перегородкой, состоящей из крупных железистых клеток, являющихся собственно „плацентой". Задняя часть образована более тонкой пленкой - производным гиподермы раковины. Поперечная перегородка имеет форму купола, вдающегося в полость выводковой сумки, по мере роста эародышей она распрямляется и прижимается к кишечнику. Выводковая сумка имеет связь с внешней средой отверстием дорсального кармана и сверху покрыта хитиновой оболочкой. Она невелика у молодых самок и разрастается по мере роста зародышей и увеличения их количества. Выводковая сумка функционирует как матка. Развитие партеногенетического яйца происходит в яичнике самки. У молодых самок яичники меньше и они имеют округлую форму. У взрослых самок яичники теряют свою правильную форму и сильно увеличиваются в размере.
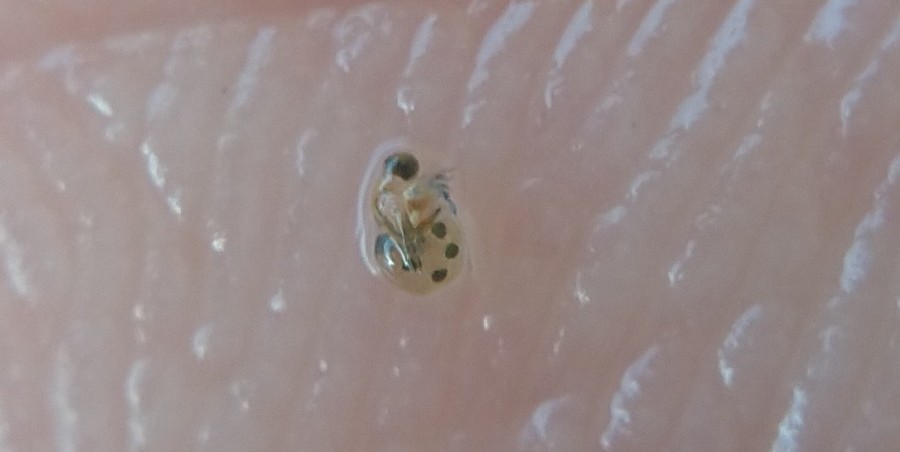 Photo © Mikko Heikkinen / iNaturalist.org. CC BY-NC 4.0
Photo © Mikko Heikkinen / iNaturalist.org. CC BY-NC 4.0
В конце мая, при низких температурах воды яйца появляются в выводковой камере на 7-11-й день жизни самки; в июле, при температуре 17-24°С - на 1-4 й день; в августе, при 17-22° - на 3-4-й день. Число яиц зависит от возраста рачков, а также от сезона. Количество яиц может достигать 40; на Рыбинском водохранилище оно колеблется от 2 до 28, постепенно увеличивается от помета к помету, но наиболее плодовиты самки весной в мае - начале июня, в июле и августе количество яиц не превышает 16. По мере старения самки плодовитость уменьшается. Продолжительность эмбрионального периода зависит в первую очередь от температуры, но также и от условий питания и разнокачественности поколений. Длительность развития первого помета в июне - июле составляет 2-6 дней, последующих - в июне - 1-2, в июле - 1-3 дня. В августе эмбриональный период удлиняется для всех пометов до 2-6 дней.
Процесс отрождения молоди происходит в несколько этапов. Рачки не имеют свободной связи выводковой сумки с внешней средой. Эмбрионы отрождаются через расширяющиеся отверстии дорсальных карманов. По мере высвобождения молодых рачков стенки выводковой сумки спадаются. Поперечная перегородка сокращается и выдавливает эмбрионы во внешнюю среду. Отрождение молоди сопровождается сбрасыванием хитиновой выводковой камеры и общей линькой рачка. Резкое сокращение числа эмбрионов предшествует началу периода обоеполого размножения.
 Photo © A. Hobæk, Norwegian Institute for Water Research. v.3 BOLD Systems. CC BY-NC-SA (2015)
Photo © A. Hobæk, Norwegian Institute for Water Research. v.3 BOLD Systems. CC BY-NC-SA (2015)
Имеет очень сложный цикл чередования партеногенетического и гамогенетического периодов. Периоды массового появления самцов и образования латентных яиц сменяются интенсивным партеногенетическим размножением. Такая полицикличность связана с чрезвычайной неустойчивостью режима мелководий, где обитает этот вид, и характерна для многих видов осушной зоны и временных водоемов. Для одного и того же вида продолжительность цикла может меняться в широтном направлении. При продвижении на юг жизненный цикл удлиняется. В течение апреля - октября отмечается несколько периодов усиления партеногенетического размножения, заканчивающихся появлением самцов, гамогенетических самок и откладкой зимних яиц. Между этими периодами рачки полностью не исчезают из водоема. Партеногенетические самки P. pediculus отрождают 3 типа пометов: чисто партеногенетические (в начале цикла), смешанные, состоящие из партеногенетических самок и гамогенетических особей обоих полов, и чисто гамогенетические, состоящие из самцов и гамогенетических самок в самых различных сочетаниях.
Самки, выпупившиеся из зимних яиц, как правило, размножаются исключительио партеногенетически, что свойственно и другим обитателям временных водоемов, например Daphnia pulex. Благодаря размножению этих особей нарастает численность популяции. В мае у P. pediculus наиболее короткий жизненный цикл, уже во втором поколении появляются самцы и гамогенетические самки. В июне увеличивается число партеногенетических поколений до 2-3, а затем следуют гамогенетические. В июле число партеногенетических поколений увеличивается до б между двумя смешанными или гамогенетическими. В августе жизненный цикл опять укорачивается, между двумя гамогенетическими поколениями появляется только одно партеногенетическое и в сентябре партеногенетически размножающиеся самки исчезают. Популяция состоит лишь из самок, производящих самцов, и гамогенетических самок, продуцирующих зимние яйца.
 Photo © CBG Photography Group, Centre for Biodiversity Genomics. v.3 BOLD Systems. CC BY-NC-SA (2011)
Photo © CBG Photography Group, Centre for Biodiversity Genomics. v.3 BOLD Systems. CC BY-NC-SA (2011)
При копуляции самец отыскивает в стае созревшую гамогенетическую самку, седлает ее и прикрепляется ногами I пары, снабженными крючками для удержания самки. Пенисы самца проникают в половые отверстия самки, и сперма попадает в яйцеводы. В семенниках у этого вида сперматозоидов бывает 12-40 в зависимости от размера самца. При прохождении через узкий канал пениса сперматозоиды благодаря своей эластичной оболочке, растягиваются, принимают веретенообразную форму, но, выходя из пениса, опять округляются. Сперматозоиды имеют темно-серый цвет и проходят по семепроводу по одному.
Гамогенетические самки способны к откладке лишь одной порции зимних яиц. Ко времени полного созревания яйца самка становится совершенно прозрачной и, по-видимому, перестает питаться: кишечник резко спадается, все ткани рачка истончаются. После созревания и откладки зимних яиц самка погибает.
К образованию зимних яиц способны только самки, родившиеся гамогенетическими. Латентные яйца тяжелые и, будучи высвобождены из раковины, в лабораторных условиях быстро погружаются на дно сосуда. Яйца имеют толстую, твердую хитиновую наружную, оболочку и под ней 2 тонких и прозрачных, внутренняя из которых окружает эмбрион. Наружная оболочка темно-коричневого цвета. Яйца требуют для своего развития длительного промерзания. При вылуплении твердая наружная оболочка яйца лопается, а самая внутренняя растягивается в виде короткого цилиндра. Через некоторое время лопается и она, и эмбрион выходит наружу. Из латентного яйца выходит молодой рачок, сходный в общих чертах со взрослыми особями. Выпупившийся из зимнего яйца P. pediculus лишен некоторых щетинок на антеннах II, а имеющиеся щетинки не оперены. Черты взрослой особи рачок приобретает только после второй линьки.
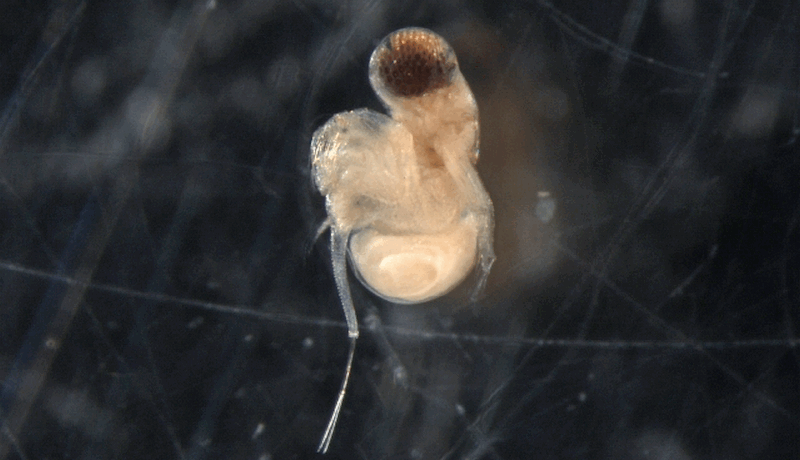 Photo © CBG Photography Group, Centre for Biodiversity Genomics. v.3 BOLD Systems. CC BY (2005)
Photo © CBG Photography Group, Centre for Biodiversity Genomics. v.3 BOLD Systems. CC BY (2005)
Ночной подъем к поверхности объясняется в значительной степени особенностями размножения - отрождением молоди. Этот процесс сложный и длительный, связанный с линькой. Кроме того, молодь питается мелкими объектами. Новорожденные появляются у поверхности, где больше коловраток, науплиев, мелких Cladocera. Количество отрождающих молодь самок изменяется в течение суток; 80% всех размножающихся за сутки особей отрождают молодь от 4 до 6 ч утра - в предрассветные и рассветные часы, днем размножение не происходит.
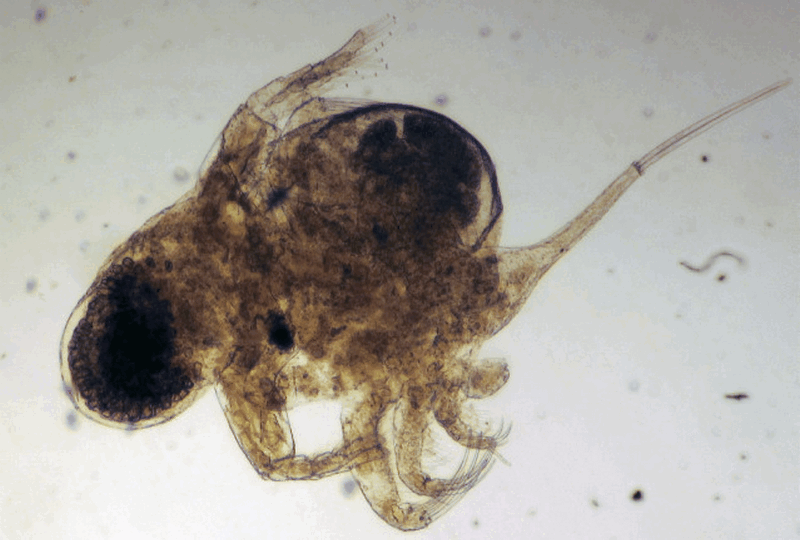 Photo © Manuel Elias Gutierrez. v.3 BOLD Systems. CC BY-NC (2009)
Photo © Manuel Elias Gutierrez. v.3 BOLD Systems. CC BY-NC (2009)
Хищник, активно ловящий добычу, с широким спектром питания - от мелких коловраток и простейших до личинок хирономид. Размер пищевых объектов 4-520 мк. Когтями ног удерживает ее, разрывает мандибулами и высасывает содержимое, жесткие хитиновые части добычи отбрасывает. Имеет хорошо развитые экзоподиты на I-III парах ног с 5-б оперенными щетинками, но никаких фильтрационных токов рачок не создает, а питается как хищник-хвататель. В лабораторных условиях у него отсутствует адаптивный период. Рачок наполняет свой кишечник за 1-2 мин. Переваривание пищи занимает всего 10-25 мин, после чего рачок снова приступает к захвату жертв. Полифем схватывает только движущуюся, окрашенную, живую добычу, ориентируясь при этом зрением. Неподвижная добыча, детрит и водоросли служат лишь дополнительным источником питания. Водоросли потребляются только голодными рачками, а усваиваются очень незначительно. Суточный прирост рачков за счет животного корма составляет 0,77-26.3%, за счет растительной пищи всего - 0,3-0,73%. Предпочитает относительно мягкую добычу. Для него недоступны покрытые твердым хитином хидориды или шиповатые ракообразные и коловратки, а также стремительные циклопы. Молодь его размером 0,2-0,35 мм ловит преимущественно простейших, беспанцирных коловраток. Самцы также питаются более мелкими животными, чем взрослые самки. Крупные самки могут поедать личинок хирономид I возраста, крупных простейших, молодь веслоногих и ветвистоусых и даже способны к каннибализму.
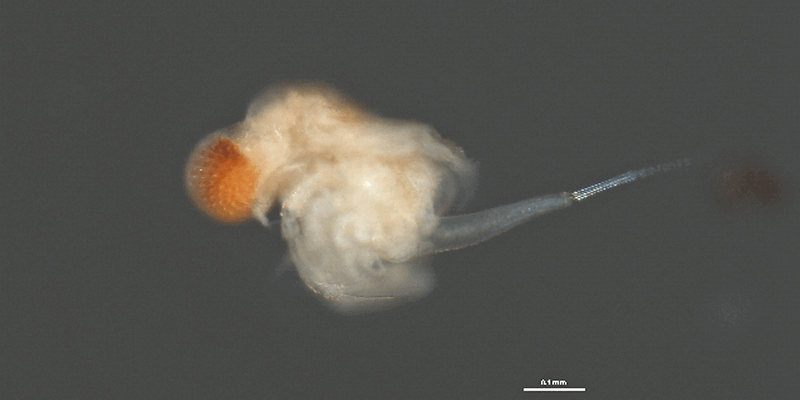 Photo © CBG Photography Group, Centre for Biodiversity Genomics. v.3 BOLD Systems. CC BY-NC-SA (2009)
Photo © CBG Photography Group, Centre for Biodiversity Genomics. v.3 BOLD Systems. CC BY-NC-SA (2009)
При ловле живой подвижной добычи, не превосходящей по скорости, хищник делает вокруг нее несколько кругов, а затем рывком схватывает. Ноги I пары делают взмах, загоняя пищу в ловчую камеру, образованную конечностями II-III пары, которые смыкаются и удерживают добычу. Жертва прокусывается мандибулами, а мягкое ее содержимое засасывается с помощью сокращений пищевода в желудок, где и происходит переваривание. Дафнию хватает за голову и начинает ее поедание со стороны глаза. Несъедобные жесткие части рачок выбрасывает. Ноги IV пары в ловле и поедании добычи не участвуют, а служат для очистки ануса.
В связи с хищным питанием, необходимостью отыскать, настигнуть добычу, схватить ее и удержать, обладает сложным быстрым маневренным движением. P. pediculus двигается мелкими скачками, но может короткое время парить. По мере развития эмбрионов движение самки постепенно замедляется. Она совершает лишь короткие пробежки, голова направлена вверх, а задний конец тела опускается вниз в зависимости от величины выводковой камеры. После отрождения молоди и линьки самки опять приобретают быстроту движений и положение тела, свойственные молодым особям. Самцы двигаются более быстро, они резко меняют направление движения, перевертываются через голову и плавают на спине.
Сезонная изменчивость выражена гораздо меньше, чем возрастная, локальная и индивидуальная. Она выражается в большей плодовитости и максимальных размерах весной партеногенетических самок. Наиболее крупные размеры самок наблюдаются весной при пониженных температурах. Это связано, видимо, с северным происхождением вида и более поздним наступлением половозрелости при низких температурах. В Рыбинском водохранилище наибольшие размеры и максимальная плодовитость наблюдаются у рачков в мае.
 Photo © Manuel Elias Gutierrez. v.3 BOLD Systems. CC BY-NC (2009)
Photo © Manuel Elias Gutierrez. v.3 BOLD Systems. CC BY-NC (2009)
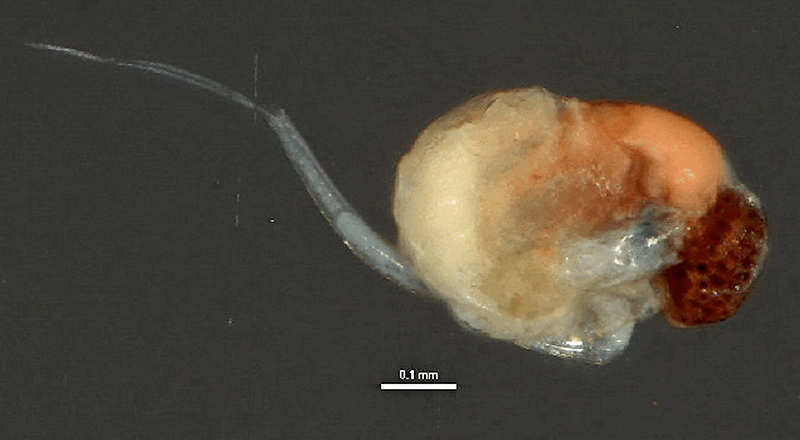 Photo © CBG Photography Group, Centre for Biodiversity Genomics. v.3 BOLD Systems. CC BY-NC-SA (2009)
Photo © CBG Photography Group, Centre for Biodiversity Genomics. v.3 BOLD Systems. CC BY-NC-SA (2009)
Литература
1. Нагорская Л. Л., Мороз М. Д., Лаенко Т. М., Вежновец В. В. "Фауна временных водоемов Беларуси". Минск, 2009. -182с.
2. Мордухай-Болтовской Ф. Д., Ривьер И. К. "Хищные ветвистоусые Podonidae, Polyphemidae, Cercopagidae и Leptodoridae фауны мира". Ленинград, 1987. -182 с.
3. Мануйлова Е. Ф. "Ветвистоусые рачки (Cladocera) фауны СССР". Москва-Ленинград, 1964. -328с.
4. Вежновец В. В. "Ракообразные (Cladocera, Copepoda) в водных экосистемах Беларуси: каталог. Определительные таблицы". Минск, 2005. -150с.