Asellus aquaticus Linnaeus, 1758 (водяной ослик)
 Photo © Ad Snijders on www.nederlandsesoorten.nl CC BY (2018)
Photo © Ad Snijders on www.nederlandsesoorten.nl CC BY (2018)
Семейство Asellidae (подотряд Asellota).
Длина около 12 мм. Тело сплющено в дорсо-вентральном направлении. Невзрачная коричневато-серая окраска водяного ослика служит ему хорошим камуфляжем и отлично гармонирует с общим тоном дна загрязненных водоемов. Как и у всех Isopoda, голова водяного ослика представляет собой типичный синцефалон, в состав которого вошел 1-й грудной сегмент. Сложная голова имеет прочный наружный хитиновый покров и не несет следов сегментации. По бокам головы расположены по четыре простых глазка. Грудной отдел слагается из 7 свободных грудных сегментов. Карапакс отсутствует. Скелет грудных сегментов состоит из спинной (тергит) и брюшной (стернит) пластинок. Брюшной отдел укорочен. В его состав входит 6 сегментов. Два передних сегмента свободны, остальные полностью сливаются между собой и полукруглым тельсоном и прикрыты сверху общей хитиновой пластинкой.
Антеннулы относительно короткие, одноветвистые. Выделяются три основных более крупных членика и более десяти мелких. Антенны имеют длину, почти равную длине тела. В их состав входят пять основных крупных члеников и множество мелких.
 Photo © Michel Renou / Galerie du Monde des insectes / www.galerie-insecte.org. CC BY-NC (2019)
Photo © Michel Renou / Galerie du Monde des insectes / www.galerie-insecte.org. CC BY-NC (2019)
Ротовое отверстие помещается на нижней стороне головы и окружено ротовыми конечностями. Спереди оно прикрыто губой - хитинизированной складкой покровов. Мандибулы мощные, состоят из основания, двух жевательных лопастей и четырехчленистого щупика. Одна из лопастей сильно хитинизирована и служит, главным образом, для измельчения пищи. Максиллы I и II нерасчлененные. Первые максиллы имеют две, а вторые три лопасти, покрытые волосками.
Первая пара грудных конечностей превращена в ногочелюсти. Они расположены на первом грудном сегменте, который вошел в состав сложной головы. Каждая ногочелюсть состоит из протоподита, включающего два членика - коксоподит и базиподит, и пятичленикового щупика. Последний, по-видимому, является эндоподитом. Базиподит покрыт волосками и несет пластинчатую лопасть. Правая и левая ногочелюсти соединены особыми волосками, расположенными по внутреннему краю базиподита. На каждой ногочелюсти имеется по два эпиподита, из которых наружный развит особенно сильно.
Свободные грудные сегменты несут по паре одноветвистых торакопод (грудных конечностей). Всего их 7 пар. Экзоподит редуцирован. Эндоподит состоит из 7 члеников: коксоподита, базиподита (самый крупный членик), ишиоподита, мероподита, карпоподита, проподита и дактилоподита. У первой пары торакопод коксоподит сливается с туловищем, и свободными остаются 6 члеников конечности. Размеры торакопод увеличиваются спереди назад. В естественном состоянии три первые пары грудных ножек расположены под углом к продольной оси тела, 4-я направлена вниз, три последние пары обращены назад.
 Photo © gailhampshire / Flickr. Upload from Wikimedia Commons. CC BY 2.0
Photo © gailhampshire / Flickr. Upload from Wikimedia Commons. CC BY 2.0
В период размножения у самок формируется выводковая камера, расположенная на брюшной стороне тела и имеющая вид зеленоватого вздутия. В ее образовании принимают участие тонкие хитиновые пластинки - оостегиты, развивающиеся в основании четырех передних пар торкопод. Тесно смыкаясь, они и формируют выводковую камеру, в которой происходит развитие яиц. Молодые особи внешне вполне похожи на взрослых. Они могут самостоятельно двигаться и через особую щель покидают выводковую сумку самки.
Брюшные конечности водяных осликов имеют специфическое строение, связанное с выполняемыми функциями. Самцы отличаются от самок строением двух передних пар абдоминальных конечностей. 1-я пара состоит из двух плоских пластинок, соединенных друг с другом с помощью хитиновых крючочков. Дистальный членик несет многочисленные перистые волоски. 2-я пара конечностей состоит из плоского базального и двух дистальных члеников, независимо прикрепленных к основному. Внутренний дистальный членик имеет крючковидный вырост. Считается, что вторая пара брюшных ножек у самцов играет важную роль при направлении семенной жидкости к половым отверстиям самок. Они функционируют как шприцы, которыми самец набирает сперму и впрыскивает ее в половые отверстия самки. 1-я пара брюшных ножек играет вспомогательную роль в этом процессе. У самок 1-я пара абдоминальных конечностей имеет вид овальной пластинки, покрытой перистыми волосками. 2-я пара отсутствует.
 Photo © Guillaume JACQUEMIN / Galerie du Monde des insectes / www.galerie-insecte.org. CC BY-NC (2019)
Photo © Guillaume JACQUEMIN / Galerie du Monde des insectes / www.galerie-insecte.org. CC BY-NC (2019)
Остальные брюшные конечности устроены у самцов и самок одинаково и выполняют дыхательную функцию. В состав этих ножек входит короткий базальный членик и две широкие листовидные пластинки, представляющие собой экзо- и эндоподит. Они тонкостенные, обильно снабжаются гемолимфой, благодаря чему выполняют функцию жабр. Пластинки расположенных рядом конечностей накладываются друг на друга и направлены таким образом, что находятся под брюшком. Экзоподит 1-й пары таких жаберных ножек имеет крупные размеры, сильно хитинизирован и делится поперечным швом на две половины. Он образует крышечку, которая прикрывает остальные жаберные ножки. Последняя пара абдоминальных конечностей является уроподами. Каждая уропода покрыта волосками и состоит из удлиненного базального членика и двух прикрепленных к нему нерасчлененных экзо- и эндоподитов. Благодаря осязательным волоскам уроподы выполняют функцию механорецепторов.

Photo © Guillaume JACQUEMIN / Galerie du Monde des insectes / www.galerie-insecte.org. CC BY-NC (2019)
 Photo © Guillaume JACQUEMIN / Galerie du Monde des insectes / www.galerie-insecte.org. CC BY-NC (2019)
Photo © Guillaume JACQUEMIN / Galerie du Monde des insectes / www.galerie-insecte.org. CC BY-NC (2019)
Самцы крупнее самок. Половому процессу предшествует длительное спаривание. Самец удерживает самку грудными ножками, располагаясь на ее спинной стороне и ожидая ее линьки (это ожидание может длиться около недели). Перенос спермы возможен только в течение 10 часов после линьки самки, когда ее половые отверстия расширены и не образовались пластинки выводковой сумки. Линька всегда происходит в два приема: сначала старый панцирь сбрасывается с задней, а потом с передней части тела. В промежуток между этими двумя линьками самец своими копулятивными брюшными ножками переносит сперму в половые пути самки. Через 2-10 часов у самки образуется выводковая сумка и туда откладываются оплодотворенные в половых путях яйца. Развивающиеся зародыш и личинка 1-й стадии обладают своеобразными боковыми придатками, функция которых не выяснена; по мнению одних исследователей это эмбриональные жабры, по мнению других - расталкиватели, с помощью которых зародыши и личинки раздвигают своих соседей, стесняющих их дыхание и питание; при первой линьке, происходящей внутри камеры, эти органы сбрасываются. Две следующие линьки также проходят внутри камеры, после чего метаморфоз водяного ослика заканчивается, и из камеры выходит вполне развитой маленький рачок (манка), длиною 1,5 мм, белого цвета. Листки, образующие выводковую камеру, подвижны: они могут то сходиться, то расходиться, причем вследствие изменения объема камеры вода то входит, то выходит из нее. Назначение этих движений - вентиляция камеры и снабжение развивающихся в ней яиц и молоди кислородом. По мере развития движение листков учащается: сначала оно совершается 1 раз в минуту, потом 2-3 раза, а перед выходом молоди до 15 раз в минуту. Камера обслуживается и другим способом: направленный внутрь её отросток ногочелюстей со щетинками ритмически колеблется, делая 2-4 удара в секунду; таким образом, камера предохраняется от засорения извне и вместе с тем удаляются экскременты молоди и сброшенные личиночные шкурки; щетинки щетки, следовательно, играют роль прочистителей камеры. Не выяснено точно, как питаются рачки внутри зародышевой камеры; известно, что стенками её выделяется голубовато-белая жидкость, которую многие исследователи склонны считать за питательную среду для (молоди ослика; но остается неясным, каким образом может происходить питание этой жидкостью в условиях постоянной проточности пульсирующей и прочищаемой камеры?
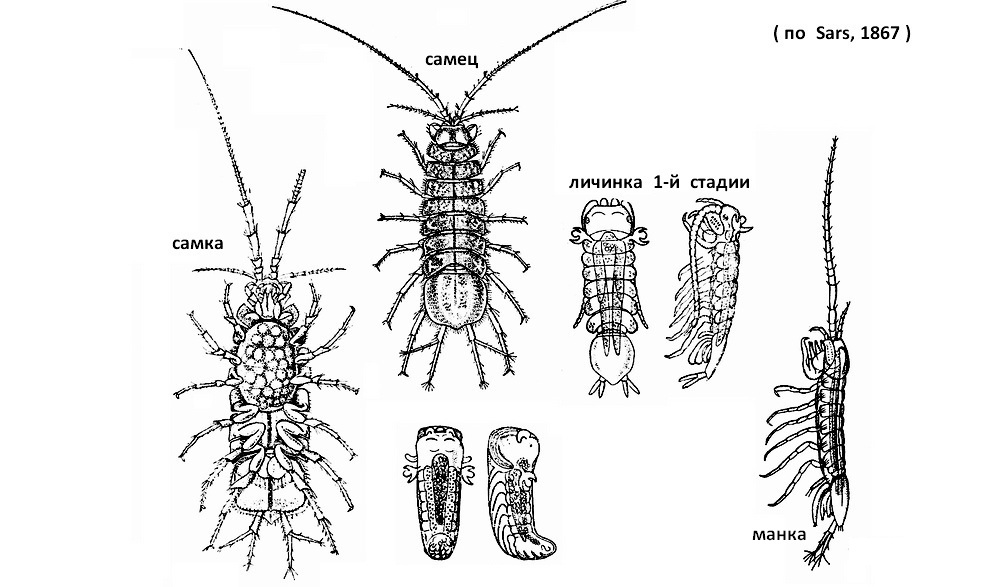
Объём выводковой камеры за время развития не меняется, размеры же развивающейся молоди увеличиваются, а число экземпляров её уменьшается. В камере одной из самок изначально было насчитано 150 яиц с эмбрионами, затем 100 личинок второй стадии, а готовых к выходу молодых осликов было лишь 80. Около половины яиц, зародышей и личинок во время развития выбрасывается через щели между листками камеры. Вытолкнутые яйца и личинки здоровы и жизнеспособны, но вне камеры развитие их прекращается.
Из выводковой сумке выходят молодые рачки, находящиеся на стадии манка. Обычно эти молодые рачки отличаются от взрослых главным образом недоразвитием последней пары грудных ножек. Самка после выхода молоди линяет, при этом выводковая сумка сбрасывается; вновь развивается она лишь после следующего спаривания и линьки.
Летом всё развитие занимает около 3 недель, зимой около 6. У водяного ослика под Москвой эмбриональное развитие длится от 14 дней в мае до 22 дней в августе. Молодые водяные ослики находят повсюду обильную пищу, быстро растут и линяют; летом того же года они дают первое потомство. Размножение продолжается осенью и весной будущего года, после чего рачок, дав очередное новое потомство, умирает. Самки водяного ослика достигают половозрелости через 50-55 дней после рождения, перелиняв за это время 13-15 раз. Живет водяной ослик 1-2 года.
 Photo László Forró / Hungarian Natural History Museum. CC BY-NC-ND 3.0
Photo László Forró / Hungarian Natural History Museum. CC BY-NC-ND 3.0
Вместе с тем, сроки развития, жизненный цикл и продолжительность жизни широко варьируются географически. Показано, что у популяций Южной Европы эмбрионы / личинки, устойчивы к 4°C, продолжительность жизни 9 месяцев, небольшой размер тела и отсутствие репродуктивной диапаузы; а у популяций Северной Европы продолжительность жизни 20 месяцев, большой размер тела, длительная репродуктивную диапауза и эмбрионы / личинки, не устойчивы к 4°C. Продолжительность жизни и репродуктивная диапауза являются генетическими признаками. Образец жизненного цикла Asellus aquaticus описан на оз. Остеншёватн в Осло. Зимующие экземпляры (генерация А) размножаются весной. Следующее поколение B созревает, размножается и умирает в первое лето, давая начало зимней генерации C, которая размножается следующей весной и умирает. Конец сезона размножения осенью, таким образом, приводит к полной смене поколений. Вместе с тем, не совсем ясно, следствием чего является подобный жизненный цикл. Объяснения выдвигается два: либо это стимуляция светом, либо предопределенный цикл, на который не влияют факторы окружающей среды.
 Photo © Michel Renou / Galerie du Monde des insectes / www.galerie-insecte.org. CC BY-NC (2019)
Photo © Michel Renou / Galerie du Monde des insectes / www.galerie-insecte.org. CC BY-NC (2019)
Размножение в оз. Остеншёватн начинается весной, когда температура воды достигает около 7-8°C. Расчетное время инкубации составляло 23–35 дней, что соответствует примерно 300 градусо-дням выше 4°C. С ноября по март-апрель роста рачков не было. Рост начинался, когда температура воды достигала 3–5°C. Увеличение роста за градусо-день выше 3°C было одинаковым для трех поколений, несмотря на разную продолжительность жизни и разные температуры. Общее количество градусо-дней выше 3°C, доступных для генераций, казалось, регулировало максимальную длину особей в соответствующих генерациях.
Широко распространенный в пресных водоемах Беларуси. Особенно часто водяные ослики встречаются в стоячих или слабопроточных водоемах, загрязненных растительными остатками. Здесь они ползают по дну. Эти ракообразные достаточно хорошо разводятся в аквариумах. Общая продолжительность жизни около 2 лет.
Типичный растительноядный. Помимо водных растений, живых и отмирающих, водяной ослик в большом количестве поедает листья деревьев, падающие в водоемы. За всю свою жизнь каждый водяной ослик потребляет 168 мг листьев (в сыром весе). В водоемах, в которые попадает много древесных листьев, водяные ослики обыкновенно живут в большом количестве.
В Беларуси A. aquaticus обнаружены как промежуточные хозяева скребней паразитов рыб Acanthocephalus anguillae, A. lucii.
 Photo © Dick Belgers on www.nederlandsesoorten.nl CC BY (2013)
Photo © Dick Belgers on www.nederlandsesoorten.nl CC BY (2013)
 Photo © Nicolas Baudet / Galerie du Monde des insectes / www.galerie-insecte.org. CC BY-NC (2019)
Photo © Nicolas Baudet / Galerie du Monde des insectes / www.galerie-insecte.org. CC BY-NC (2019)
Литература
1. Круглова О. Ю. "Класс Crustacea – Ракообразные : метод. рекомендации". В 2 ч. Ч. 2. Высшие ракообразные. Минск, 2014. –43с.
2. Бирштейн Я. А., Пастернак Р. К. "Отряд Равноногие ракообразные (Isopoda)" / Жизнь животных. 2-е изд. Т. 2. Моллюски. Иглокожие. Погонофоры. Щетинкочелюстные. Иглокожие. Полухордовые. Хордовые. Членистоногие. Ракообразные. Москва, 1988. С.365-377
3. Голубеў А. П. "Раўнаногія ракападобныя" / Энцыклапедыя прыроды Беларусі. У 5-i т. Т. 4. Недалька - Стаўраліт. Мінск, 1985. С.370-371
4. "Макрыцы (Oniscoidea)" / Беларуская энцыклапедыя: У 18 т. Т. 9: Кулібін - Малаіта. Мінск, 1999. С.543
5. Павловский Е. Н., Лепнева С. Г. "Очерки из жизни пресноводных животных". Ленинград,, 1948. -458с.
6. Vitagliano G., Fano E. A., Marchetti E., Colangelo M. A., Vitagliano E. (1991) Importance of longevity, growth, and diapause in the evolution of Asellus aquaticus / Bolletino di zoologia, 3: 125–131.
7. Økland, K.A. (1978) Life history and growth of Asellus aquaticus (L.) in relation to environment in a eutrophic lake in Norway / Hydrobiologia. Volume 59, Issue 3, pp.243-259
8. Бычкова Е. И., Акимова Л. Н., Дегтярик С. М., Якович М. М. Гельминты позвоночных животных и человека на территории Беларуси. Каталог. Минск, 2017. -316с.