Bythotrephes longimanus Leydig, 1860
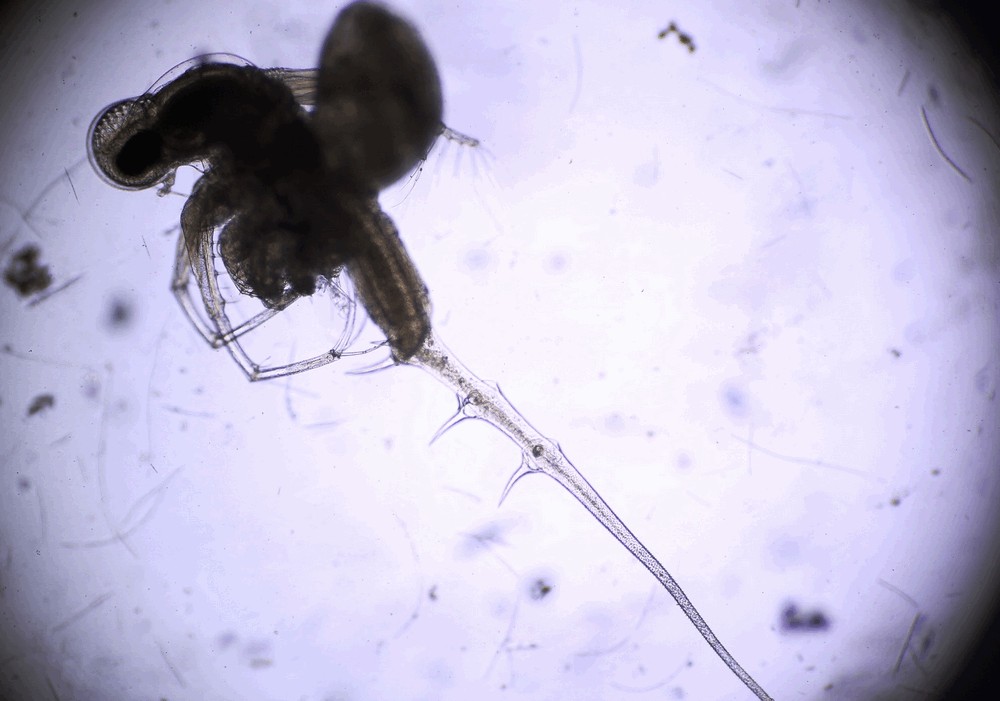 Photo © Надежда (Надежда Климова) / iNaturalist.org. CC BY-NC 4.0
Photo © Надежда (Надежда Климова) / iNaturalist.org. CC BY-NC 4.0
Onychopoda. Семейство Cercopagididae.
В Беларуси распространен преимущественно в ее северной части. Обитатель многих озер.
Тело прозрачное. голубоватое или желто-красное. Длина (без хвостовой иглы) самки 2,0-5,0, самца 1,8-4,0 мм. Туловище хорошо подразделяется на отдельные части: голову, грудные ноги, выводковую сумку, абдомен, кауду с хвостовым придатком. Глазной отдел головы шаровидный, целиком занят крупным сложным глазом. Ротовой отдел несет верхнюю губу, за которой расположены мандибулы. Грудных ног четыре пары. Они одноветвисты, членики узкие и длинные. Пара I значительно длиннее остальных трех пар. Плавательные антенны относительно большие, 4-члениковая ветвь несет 8 щетинок, 3-члениковая - 7. Абдомен цилиндрической формы, несколько выпуклой спереди, состоит из двух члеников - короткого и длинного. Последний имеет глубокую хитиновую складку с дорсальной стороны. Выводковая сумка служит лишь для вынашивания зародышей и растет по мере их развития. Кауда в виде бугорка с когтями, между которыми открывается анальное отверстие. Сзади кауда продолжается в длинную хвостовую иглу с двумя плавательными щетинками на конце.
 Photo © Spencer Hardy / iNaturalist.org. CC BY-NC 4.0
Photo © Spencer Hardy / iNaturalist.org. CC BY-NC 4.0
Партеногенетическая самка. В передней шаровидной части головного отдела - голове помещается крупный, сильно пигментированный глаз. За ним находится небольшая выемка, в которую открываются протоки железистого органа. Головной щит сильно развит, выпуклый, через его покровы просвечивают пигментированные 5-6-гранные железистые клетки. За ним следует небольшой хитиновый валик, отделяющий головной отдел от туловища. Внутри этой выпуклой части прикрепляются сильные мышцы, приводящие в движение мандибулы. Головной отдел подразделяется на две части: переднюю - церебральную, где находится глаз, головной ганглий, прикрепляются антенны, и заднюю - мандибулярную. Глазной пигмент в виде полукруглой массы расположен в передне-нижней части глаза, от него во все стороны отходят многочисленные фасетки - омматидии (около 300). Омматидии имеют форму конуса и разделены между собой прослойками пигмента. Движение глаза обеспечивается тремя парами мускулов. Пигментное пятно, или науплиев глаз, отсутствует. Антеннулы небольшие, грушевидной формы, находятся на нижней части глазного отдела головы. Они короткие, одночлениковые, отходят от общего основания, обладают слабой подвижностью. Каждая антеннула на конце несет 5 эстетасков - тонкостенных папилл - органов химического чувства. Кроме папилл антеннулы несут чувствительную щетинку, выполняющую осязательную функцию. Плавательные антенны прикрепляются к задней части головного отдела складчатым основанием протоподита. Складчатый отдел короткий, протоподит (ствол) цилиндрический, толстый, наполненный мышцами. Наружная четырехчлениковая ветвь антенны несет 8 щетинок, внутренняя трехчлениковая - 7. Плавательные щетинки двучлениковые. Нижняя часть имеет более мягкое оперение, дистальная - жесткое. Ротовые части состоят из верхней губы, мандибул и максилл I, или максиллул. Верхняя губа крупная, подвижная, прикрывает собой почти весь комплекс ротовых частей. На передне-верхней стороне губа несет большой серпообразный вырост, конец которого заходит ниже края губы. Вырост покрыт рядами шипиков, край губы - рядом более мелких шипиков. Сверху губа имеет ромбовидную форму, на переднем углу губы располагаются 5-6 более крупных шипиков. У основания губы непосредственно перед ротовым отверстием хорошо видны заостренные концы мандибул. Мандибула имеет изогнутую форму, основание ее в виде широкой пластины. Сверху концы мандибул представляют собой трехзубую вилку. Снизу под зубцами располагается округлый выступ, несущий б-8 папилл и одну тонкую, видимо, осязательную щетинку. На нижней стороне мандибулы имеется ряд оперенных щетинок. Маленькие максиллы I представляют собой округлые пластинки, опушенные по краям. На вершине максиллулы несут по 3 трубкообразных папиллы и одну осязательную щетинку.
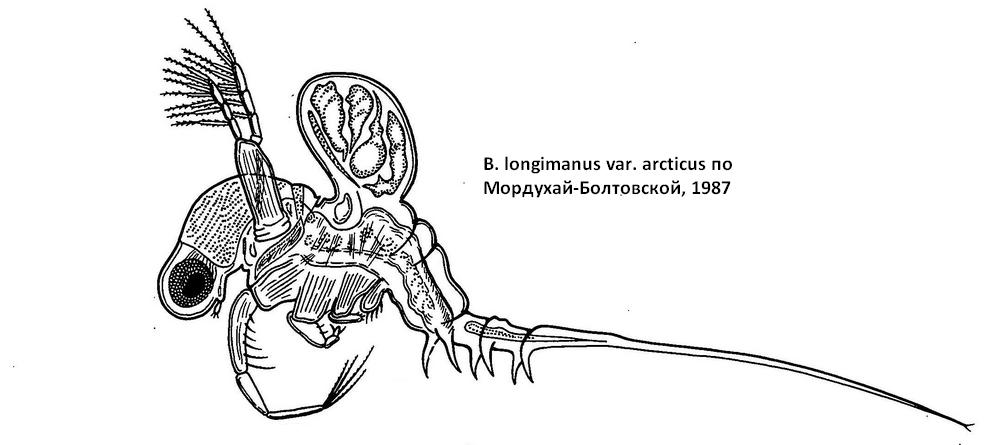 Туловище короткое, широкое, спереди несет 4 пары конечностей, сзади - выводковую сумку. Конечности одноветвистые, палочковидные; I пара длиннее остальных, IV заметно редуцирована. Членики соединены суставами. С внутренней стороны I-IV пары снабжены максиллярными придатками, с наружной I-III пары ног имеют шипообразной формы выросты, не отчлененные от базиподита. Максиллярный придаток на I паре в виде треугольного выроста с шипиками, коническим отростком и осязательной щетинкой. На II-IV паре максиллярные придатки направлены внутрь и представляют собой пальцеобразные выросты с шипами по верхнему краю. Первые членики I-III пары несут по 4-10 оперенных щетинок. Дистальные края первого и второго члеников несут парные щетинки, из которых одна мягкая, короткая, оперенная, а другая длинная, гладкая, нитевидная. Третий дистальный членик I-III пары несет по 4 мощных, покрытых рядами шипиков, когтевидные щетинки, из которых одна заметно длиннее остальных, а три другие снабжены крючочками на концах. IV пара конечностей имеет укороченные, почти слитые членики, однако общее количество щетинок такое же, как на остальных конечностях, три концевые щетинки также снабжены крючочками. С вентральной стороны между конечностями тело образует желоб. Задняя сторона туловища несет выводковую сумку, зачаточную у молодых рачков и самцов. При большом количестве зрелых зародышей выводковая сумка разрастается и превосходит размеры тела в 2-3 раза. Внутренняя оболочка выводковой сумки с дорсального края состоит из крупных железистых клеток. Брюшко цилиндрической формы имеет на поверхности 2-3 поперечных складки, слабо сегментировано. Кишечник не образует петли. Анальное отверстие лежит на конце постабдомена между двумя коготками. Каудально постабдомен образует вырост - хвостовой отросток, на котором по мере роста рачка образуется 3-4 пары когтей. Эти когти представляют собой оставшиеся после линек хитиновые футляры хвостового стебля со старыми когтями, отодвинутыми вновь появившимися кзади. У молодых особей на конце хвостового придатка имеются две маленькие „плавательные" щетинки, у взрослых особей они, видимо, утрачиваются. Каудальные когти бывают различных размеров и направлены или вперед, или назад. При наличии длинных, загнутых вперед когтей хвостовой придаток имеет волнообразный изгиб, покрытый крепкими, загнутыми шипиками.
Туловище короткое, широкое, спереди несет 4 пары конечностей, сзади - выводковую сумку. Конечности одноветвистые, палочковидные; I пара длиннее остальных, IV заметно редуцирована. Членики соединены суставами. С внутренней стороны I-IV пары снабжены максиллярными придатками, с наружной I-III пары ног имеют шипообразной формы выросты, не отчлененные от базиподита. Максиллярный придаток на I паре в виде треугольного выроста с шипиками, коническим отростком и осязательной щетинкой. На II-IV паре максиллярные придатки направлены внутрь и представляют собой пальцеобразные выросты с шипами по верхнему краю. Первые членики I-III пары несут по 4-10 оперенных щетинок. Дистальные края первого и второго члеников несут парные щетинки, из которых одна мягкая, короткая, оперенная, а другая длинная, гладкая, нитевидная. Третий дистальный членик I-III пары несет по 4 мощных, покрытых рядами шипиков, когтевидные щетинки, из которых одна заметно длиннее остальных, а три другие снабжены крючочками на концах. IV пара конечностей имеет укороченные, почти слитые членики, однако общее количество щетинок такое же, как на остальных конечностях, три концевые щетинки также снабжены крючочками. С вентральной стороны между конечностями тело образует желоб. Задняя сторона туловища несет выводковую сумку, зачаточную у молодых рачков и самцов. При большом количестве зрелых зародышей выводковая сумка разрастается и превосходит размеры тела в 2-3 раза. Внутренняя оболочка выводковой сумки с дорсального края состоит из крупных железистых клеток. Брюшко цилиндрической формы имеет на поверхности 2-3 поперечных складки, слабо сегментировано. Кишечник не образует петли. Анальное отверстие лежит на конце постабдомена между двумя коготками. Каудально постабдомен образует вырост - хвостовой отросток, на котором по мере роста рачка образуется 3-4 пары когтей. Эти когти представляют собой оставшиеся после линек хитиновые футляры хвостового стебля со старыми когтями, отодвинутыми вновь появившимися кзади. У молодых особей на конце хвостового придатка имеются две маленькие „плавательные" щетинки, у взрослых особей они, видимо, утрачиваются. Каудальные когти бывают различных размеров и направлены или вперед, или назад. При наличии длинных, загнутых вперед когтей хвостовой придаток имеет волнообразный изгиб, покрытый крепкими, загнутыми шипиками.
 Photo © A. Hobæk, Norwegian Institute for Water Research. v.3 BOLD Systems. CC BY-NC-SA (2015)
Photo © A. Hobæk, Norwegian Institute for Water Research. v.3 BOLD Systems. CC BY-NC-SA (2015)
Самки делятся на партено- и гамогенетических, последние значительно крупнее первых. Гамогенетическая самка продуцирует только зимние яйца. Каудально, снизу у основания выводковой сумки имеется половое отверстие, ведущее внутрь ее, где оно открывается рядом с выходом яйцеводов. Латентные яйца в количестве до 9, крупные, желтоватого цвета, с утолщенными оболочками. Глаз у взрослых особей черно-коричневого цвета. Нижняя часть брюшка и выводковой сумки имеет синевато-фиолетовый цвет. Кишечник обычно окрашен в зеленовато-бурые тона. Длина самок без хвостового стебля 1,7-6,6 мм, длина хвостового стебля до 7 мм.
Самец. Отличается более стройным телом. Несколько меньше самки, на месте выводковой сумки имеет выпуклый рудимент, где помещается сердце. Голова и глаз относительно крупные. Глаз у самца больше, чем у самки, его диаметр составляет 22-24% длины тела; у самки - 18-20%. Кроме того, самцы имеют только две пары когтей на хвостовом придатке, в то время как у самок их 3-4. Видимо, самцы проходят на 1-2 ростовых линьки меньше, чем самки. Ноги I пары значительно короче и толще, чем у самки. У основания дистального членика I пары имеется гладкий крючок, вокруг которого поверхность членика уплотненная и шероховатая. Он служит для удерживания самки. Семепроводы открываются на концах небольших пенисов за IV парой ног. Сперматозоиды Bythotrephes представляют собой крупные малоподвижные округлые клетки. В семенниках у этого вида их бывает до 20, но чаще 6-8. При прохождении через узкий канал пениса сперматозоиды благодаря своей эластичной оболочке, растягиваются, принимают веретенообразную форму, но, выходя из пениса, опять округляются. Длина самца 1,6-5,0 мм, длина хвостового стебля до 7 мм.
 Photo CBG Photography Group on BoldSystems v.4. CC BY-NC-SA
Photo CBG Photography Group on BoldSystems v.4. CC BY-NC-SA
Органы дыхания устроены иначе, чем у прочих ветвистоусых. Конечности хищных форм лишены эпиподитов - дыхательных отростков или жабр. Газообмен совершается исключительно с помощью затылочного органа (так называемого головного щита), представляющего собой скопление крупных железистых клеток. Он в виде большого вздутия располагается за головой выше прикрепления плавательных антенн и примыкает со спинной стороны к сердцу. Здесь наблюдается наибольшая проницаемость покровов, происходят поглощение кислорода и выделение углекислоты. Способ дыхания посредством головного щита - единственный.
Кровеносная система устроена примитивно и состоит из сердца, имеющего пару остий (отверстий), через которые протекает гемолимфа, циркулирующая затем по телу среди мембран и перекладин внутреннего скелета. Сердце представляет собой мешочек, расположенный позади затылочного органа, непосредственно над выводковой сумкой. От сердца отходит небольшой участок развитого сосуда.
Типичные планктонные животные. Молодь сразу после рождения ведет планктонный образ жизни. Лишь одна стадия жизненного цикла - латентное (покоящееся) яйцо опускается на дно либо в глубинные слои и находится там до выхода молодого рачка.
Типичный обитатель пелагиали. Длинная, снабженная когтями хвостовая игла способствует парению рачка в толще воды. Вероятно, хвостовой придаток такого строения неудобен на заросших мелководьях, где он цепляется за водную растительность. Заселяет глубокие участки крупных водоемов, преимущественно озер и озеровидных плесов водохранилищ. Как активный мигрант он избегает и открытых незаросших мелководий, где невозможны вертикальные перемещения вглубь. Это явление, называемое „бегством от берега", видимо, связано с восприятием преломленного и отраженного от берега света или с сильным освещением до дна на мелководьях, откуда рачок с наступлением рассвета старается уйти в более глубокие, слабо освещенные участки, избегая поедания рыбами.
Летние, преимущественно теплолюбивые формы. Появление первого поколения из латентных яиц прослежено как в природе, так и в эксперименте. В лаборатории при повышении температуры до 10-12°С через 3 дня оболочки яйца лопаются и молодой рачок выклевывается. Максимальной численности в водоемах Средней полосы В. longimanus достигает в самое теплое время года при 20-24°С. Интенсивность питания снижается после 25°. Исчезает В. longimanus при значительно более низких температурах, чем появляется; при 5° в середине октября в Рыбинском водохранилище встречаются еще последние особи. В крупных озерах и водохранилищах наибольшая численность В. longimanus отмечается в устьях впадающих в водоем рек, где количество мелких ракообразных - его пищи - обычно выше. При образовании водохранилищ В. longimanus становится в них широко распространенной, обычной формой. Однако в верхних речных участках искусственных водоемов озерного и в водохранилищах руслового типа количество рачка невелико. В приплотинных расширениях, где складывается режим, близкий к озерному, численность В. longimanus возрастает в 50-100 раз. Несмотря на явную приуроченность вида к открытым водным пространствам, известно нахождение рачка в небольших арктических водоемах. Рачок легко культивируется в мелководных прудах, где всегда заселяет участки, удаленные от зарослей водной растительности.
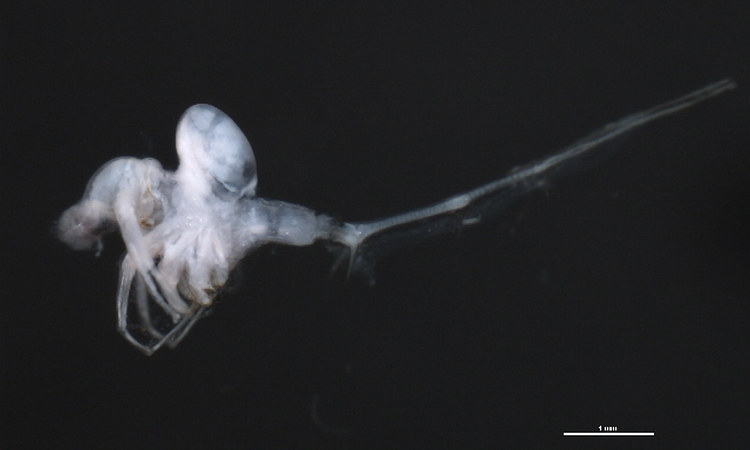 Photo CBG Photography Group on BoldSystems v.4. CC BY-NC-SA
Photo CBG Photography Group on BoldSystems v.4. CC BY-NC-SA
В глубоких озерах распределяется до термоклина; в этом же интервале и происходят его вертикальные перемещения. В наиболее глубоких приплотинных участках водохранилищ вертикальные перемещения битотрефа охватывают всю толщу воды, так как стратификация здесь неустойчива. Днем рачки держатся вблизи дна, а ночью всплывают к поверхности. Амплитуда вертикальных перемещений молодых рачков гораздо уже.
Населяет пресные водоемы самой различной степени минерализации: 50-60 мг/л - Ладожское, Онежское озера, до 400 мг/л - Цимлянское водохранилище. Имеются указания, что В. longimunus с длинным хвостовым придатком (форма cederstraemi) чаще встречается в мезо- и полигумозных, бедных Са водах. Случаи нахождения в соленых водах неизвестны. Также быстро погибают при сильной проточности.
Динамика численности в течение лета не испытывает таких резких колебаний. Это связано со значительной стабильностью условий в пелагиали, где живет этот вид. В Рыбинском водохранилище первые особи из зимних яиц выпупляются в начале мая, во второй половине июня численность возрастает и достигает максимальных величин в конце июля. Самцы появляются в популяциях на 1-1,5 мес раньше гамогенетических самок, последние отмечаются при достижении популяциями наивысшей численности - в конце июля - начале августа. Спад численности происходит более резко, чем подъем, и уже к концу августа - началу сентября количество рачков снижается в 4-5 раз (со 150 до 20 экз./м³). Заканчивают свой жизненный цикл популяции В. longimanus, имея в своем составе партеногенетических самок, самцов и гамогенетических самок с латентными яйцами. Последние, единичные особи В. longimanus держатся в водохранилище до середины октября.
 Photo © A. Hobæk, Norwegian Institute for Water Research. v.3 BOLD Systems. CC BY-NC-SA (2015)
Photo © A. Hobæk, Norwegian Institute for Water Research. v.3 BOLD Systems. CC BY-NC-SA (2015)
Вертикальные суточные перемещения довольно сложные. Утром и днем рачки находятся в средних и придонных слоях воды, а вечером и ночью поднимаются к поверхности. С 4 ч утра начинается отход рачков от поверхности и погружение их вглубь. Миграции молоди происходят иначе, чем взрослых особей, они раньше поднимаются к поверхности и раньше опускаются вглубь. В мелководных озерах миграции выражены слабее, чем в глубоких участках водохранилищ - затопленных руслах рек. Основная масса рачков в течение дня держится в среднем слое, поднимаясь к поверхности в самое темное время суток. Совсем отсутствуют миграции в прудах с глубиной 1,5-2 м, где днем рачки скапливаются в затененных участках. В глубоких водоемах рачки в течение суток находятся почти в постоянном движении. Днем они держатся над грунтом, в 5-10-сантиметровом слое, вечером поднимаются к поверхности воды, во вторую половину ночи их здесь наибольшее количество, а с рассветом начинается погружение вглубь. Наиболее четко выражены миграции летом в тихую погоду. Подъем животных к поверхности вечером длится 8-10 ч, с рассветом начинается их медленное погружение. Осенью подъем и погружение идут с большей скоростью, чем летом, и длятся всего 4-6 ч. Иногда в сентябре отмечаются два подъема рачков к поверхности - небольшой по численности (в основном взрослые самки) сразу после захода солнца, в 21 ч, и более массовый - ночью. Летом, в период коротких ночей, пребывание битотрефа у поверхности непродолжительно. Осенью, когда ночи удлиняются, сумерки кратковременны, ускоряется передвижение рачков к поверхности, где они остаются дольше, чем летом.
Новорожденные не совершают активных перемещений. Появляясь на свет ночью в поверхностном слое, новорожденные к утру начинают опускаться вглубь, но обычно никогда не достигают придонного слоя, а распределяются в толще воды (от 3 до 6 м глубины). Молодые рачки с эмбрионами на ранних стадиях развития в светлое время находятся в среднем и придонном горизонтах. Подъем их к поверхности ночью происходит медленнее, чем взрослых рачков. Общая картина вертикального распределения битотрефа в различные часы суток определяется ходом миграций взрослых особей, у которых способность к активному движению выражена в наибольшей степени. Миграции самцов и гамогенетических самок совпадают и идут параллельно с миграциями взрослых партеногенетических самок. Вертикальные перемещения В. longimanus в течение суток связаны с миграциями его кормовых объектов - ветвистоусых рачков, с уходом битотрефа от хищников - рыб, а также ритмикой размножения. Отрождение молоди у В. longimanus происходит в самое темное время суток у поверхности воды. Этим обеспечивается наибольшая сохранность нового поколения.
 Photo CBG Photography Group on BoldSystems v.4. CC BY-NC-SA
Photo CBG Photography Group on BoldSystems v.4. CC BY-NC-SA
Обладает, как все Cladocera, двумя способами размножения: партеногенетическим и гамогенетическим, с участием самцов и гамогенетических самок. Партеногенетическое размножение продолжается в течение всего жизненного цикла вида, до исчезновения последнего из водоема и является способом интенсивного наращивания численности популяцией. Гамогенез прослеживается в конце цикла; в результате: двуполого размножения появляются латентные яйца, способные переносить неблагоприятные условия.
Как и у остальных ветвистоусых, новорожденный рачок появляется на свет крупным, полностью развитым. Развитие яйца происходит в замкнутой выводковой сумке, стенки которой постепенно выделяют питательные вещества, необходимые для полного развития зародыша. Выводковая сумка функционирует как матка. Дорсальная сторона выводковой сумки состоит из крупных железистых клеток и образует „плаценту", прогибающуюся внутрь самки. Остальная часть выводковой сумки представляет собой тонкую оболочку, срастающуюся с гиподермой хитиновой камеры. Поэтому рост хитиновой, выводковой камеры происходит в строгом соответствии с ростом выводковой сумки, т. е. с ростом зародышей. Выводковая сумка сообщается тонким канальцем лишь с полостью хитиновой камеры, служащей лишь для вынашивания зародышей. Развитие партеногенетического яйца происходит в яичнике самки. У молодых самок яичники меньше и они имеют вытянутую форму. У взрослых самок яичники теряют свою правильную форму и сильно увеличиваются в размере. Количество яиц обычно колеблется от 4 до 14. Однако самки первого поколения, вышедшие из зимних яиц, несут максимальное число яиц - до 24, такая плодовитость никогда не наблюдается у следующих поколений.
Продолжительность эмбрионального периода зависит в первую очередь от температуры, но также и от условий питания и разнокачественности поколений. В природе поступление яиц у B. longimanus наблюдается летом на 2-3 сутки после рождения молодых рачков. В лабораторных условиях при одинаковой интенсивности кормления созревание партеногенетических особей зависит от температуры: в диапазоне 10-25°С сроки созревании сокращаются от 23 до 7 дней, а продолжительность периода между приплодами соответственно от 11 до 4 дней.
Процесс отрождения молоди происходит в два этапа. Сначала полностью сформированные рачки проходят под гиподерму и размещаются непосредственно в хитиновой камере, где остаются еще некоторое время. В это время плацента и вся выводковая сумка с примыкающей к ней гиподермой сокращаются. В выводковую сумку быстро перетекают яйца нового помета. Этот этап длится у В. longimanus 30-40 мин. Затем наступает второй этап отрождения молоди - выход ее во внешнюю среду, сопровождающийся разрывом хитиновой камеры. Обрывки старой хитиновой выводковой камеры иногда некоторое время держатся на спине матери, а затем отпадают. По мере роста зародыши у церкопагид приобретают способность двигаться. Особенно подвижными становятся эмбрионы после высвобождения из выводковой сумки, когда они попадают непосредственно под хитин выводковой камеры. Это помогает им быстро выскакивать на свободу. Новорожденные рачки имеют полностью сформированные конечности, пигментированный глаз, хвостовой придаток у них мягкий и разнообразно изогнутый. Через 30-50 мин хвостовой придаток твердеет и выпрямляется. У материнской особи после отрождения молодых выводковая камера очень невелика. Она треугольная, в ней видны яйца нового помета. К осени число эмбрионов сокращается до 1-3. Резкое сокращение числа эмбрионов предшествует началу периода обоеполого размножения.
Единичные самцы появляются в водоеме в начале развития популяции. Так, в Рыбинском водохранилище самцы появляются в начале июня, в третьем поколении. Самки, вышедшие из зимних яиц, размножаются исключительно партеногенетически. Первые гамогенетические самки появляются на 1,5 мес позже первых самцов, в конце июля, когда популяция достигает своей максимальной численности. В это время самцов уже в 3 раза больше, чем самок. В начале сентября основу популяции составляют гамогенетические особи, причем соотношение самцов и гамогенетических самок составляет 3:1. B. longimanus заканчивает жизненный цикл, имея в составе своей популяции, кроме самцов и самок с зимними яйцами, некоторое количество партеногенетических особей, продолжающих размножаться. У вида имеется только один период гамогенетического размножения.
Самец постоянно прикрепляется одинаково часто как к партеногенетическим, так и к гамогенетическим самкам, как это наблюдается и у многих других Cladocera. Самец совершает как бы „охотничий" бросок и седлает самку, прикрепляясь к ней сверху, однако быстро отпускает самок, не пригодных к оплодотворению, ощупывая их, видимо, жгутиком антенн I. Самец удерживает самку ногами I пары, снабженными крючками, затем посредством пенисов через половые отверстия самки сперматозоиды проникают в яйцеводы, где происходит оплодотворение. Весь процесс копуляции длится около 30-60 с. Через некоторое время (10-15 мин) яйца перетекают в выводковую сумку. Вероятно, процесс копуляции способствует перетеканию яиц. В выводковой сумке проиcходит формирование оболочек и начинается поверхностное дробление яйца.
К образованию зимних яиц способны только самки, родившиеся гамогенетическими. В отличие от P. pedicutus y B. longimanus может образоваться несколько порций зимних яиц. Откладка латентных яиц происходит посредством линьки выводковой камеры, как это наблюдается и при отрождении партеногенетических зародышей. Диаметр зрелого яйца 0,46-0,66 мм. Яйца имеют толстую, твердую хитиновую наружную, оболочку и под ней 2 тонких и прозрачных, внутренняя из которых окружает эмбрион. Наружная оболочка желтоватого цвета. Яйца требуют для своего развития кратковременного промерзания. После кратковременного промерзания, а затем помещения яиц в воду с температурой 10-12°С твердые оболочки яйца лопаются, образуя две неравные полусферы, которые отодвигаются друг от друга. Между ними несколько дней существует „манжет" из одной тонкой эластичной внутренней оболочки. Внутри увеличившегося таким образом пространства яйца шевелится эмбрион. Через 6-7 дней и эта оболочка разрывается, рачок выходит во внешнюю среду. Он имеет мягкий нитевидный хвостовой придаток с небольшими бугорками на месте коготков. Через 5-7 мин хвостовой придаток твердеет, а через 0,5 ч рачок линяет. У него появляются коготки на хвостовом придатке и недостающие щетинки на 2 и 3-м члениках I пары ног, вооружение плавательных антенн становится таким же, как у взрослого рачка. Однако молодой рачок, вышедший из латентного яйца, отличается морфологически от новорожденных последующих поколений: у него более короткий хвост, очень короткая I пара ног, отсутствует 8-образный изгиб на хвостовом придатке. В дальнейшем самки первого поколения имеют четыре пары когтей на хвостовом придатке, т. е. проходят на одну ростовую линьку больше, что резко отличает их от всех последующих поколений.
Отрождение молоди ночью тесно связано с миграциями, и в мелких водоемах (прудах) четкой ритмики в размножении не наблюдается. Летом, в период наиболее интенсивного размножения, отрождение молоди начинается в июле в 21, а в августе - в 19 ч, наибольшее количество размножающихся самок отмечается в 23 ч, процесс заканчивается в предрассветные часы - 3 ч.
Хищник. Паря в воде и выслеживая добычу, активно ловит ее. Нападает почти на всех планктонных животных, включая собственную молодь, но не трогает сидячих и даже временно прикрепляющихся рачков, например Sida и Simocephalus. Питаться начинает сразу после рождения, предпочитая мелкую и ярко окрашенную движущуюся добычу. Излюбленный корм - полифемы, босмины, цериодафнии. В первые сутки жизни В. longimanus съедает от 1 до 17 мелких рачков, размером 0,3-0,4 мм, на 5-е сутки - до 21 рачка. Суточный рацион с повышением температуры возрастает, но при 25-27°С В. longimanus перестают питаться. Взрослый рачок в эксперименте поедает в сутки до 40 рачков размером 0.6-0.7 мм. В выслеживании и поимке добычи ориентируется в основном зрением. Хищник нагоняет жертву, затем сильным броском сближается с ней, выбрасывая длинные ноги I пары. Жертва удерживается конечностями II-III пары, разрывается мандибулами и жидкое содержимое высасывается. В эксперименте взрослые самки хватают и пожирают свою молодь. Остатки жертвы отбрасываются. Эффективность охоты В. longimanus велика и составляет 80-100%. Всегда в первую очередь прокусывает голову и высасывает содержимое вместе с черным глазным пигментом.
В связи с хищным питанием, необходимостью отыскать, настигнуть добычу, схватить ее и удержать, обладает сложным быстрым маневренным движением. За компактным телом следует тонкий длинный хвостовой придаток, который служит как для увеличения площади парения, так и для создания прямолинейности движения - „устойчивости на курсе". При прямолинейном движении вперед тело составляет с хвостовым придатком почти прямую линию. Число взмахов антеннами в среднем 147 в минуту. Средняя скорость движения рачка составляет около 0,7 см/с, максимальная - 48 м/ч. Молодь рачка с одной или двумя парами когтей на хвостовом придатке движется со скоростью 0,7-0,8 см/с, тогда как скорость взрослых самок, а особенно самцов, значительно выше и составляет соответственно 0,9-1,1 см/с и 1,2-1,8 см/с. Во время охотничьего броска рачок действует молниеносно, развивая скорость 2-2,5 см/с. Скорость движения в различные моменты питания - при поиске добычи (крейсерская), погоне за ней (максимальная), в момент броска для поимки жертвы (бросковая) и в процессе поедания добычи - заметно отличаются. Скорость в момент броска возрастает в 5-7 раз по сравнению с „крейсерской".
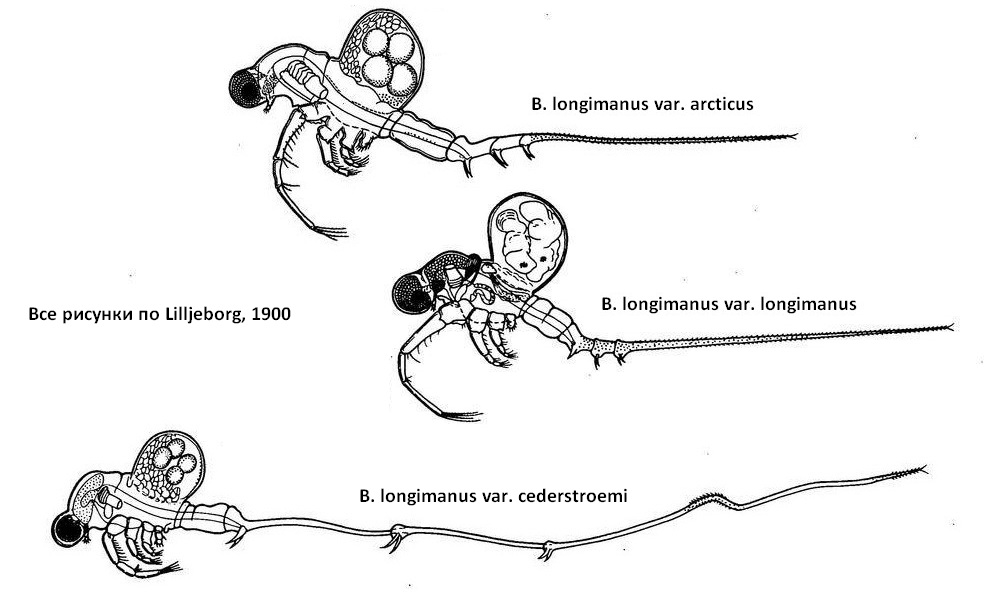
Сезонная изменчивость выражена гораздо меньше, чем возрастная, локальная и индивидуальная. Выражается в большей плодовитости и максимальных размерах весной партеногенетических самок. Вместе с тем, у B. longimanus, отмечается чрезвычайная полиморфность. Этот вид по многим источникам насчитывает 6-9 вариететов или форм. Этот длинный ряд вариететов имеет на одном конце форму В. longimanus var. arcticus - очень крупньх массивных (5,5 мм) короткохвостых рачков. Хвостовой придаток у этих форм равен длине тела и имеет сближенные маленькие коготки. На другом конце ряда находится форма В. cederstroemii var. cederstroemii (= В. longimanus cederstroemii) - мелкие рачки с легким телом длиной 1,7-2,5 мм и чрезвычайно длинным, в 3-4,2 раза длиннее тела, хвостовым придатком с хорошо выраженным изгибом с длинными, широко отодвинутыми когтями.
В Рыбинском водохранилище в течение сезона встречаются рачки с крупным телом, укороченным хвостовым придатками, без изгиба, с мелкими, близко расположенными коготками (var. longimanus) и рачки с легким телом и длинным хвостовым придатком (var. cederstroemii). При изучении встречаемости этих форм от весны к осени удалось, выявить сезонную изменчивость B. longimanus. Первое поколение этого вида, появляющееся из зимних яиц, обладает маасивным телом (2,32 мм), коротким хвостовым придатком (всего 1,71 от длины тела), лишенным изгиба, с маленькими близко расположенными коготками. Эта весенняя форма (var. longimanus) наиболее близка к тундровой var. arcticus. Постепенно, с пргреванием воды длина тела уменьшается (1,92 мм), хвостовой придаток удлиняется, становится в 3 раза длиннее тела. Длина коготков увеличивается по сравнению с весной в 4-5 раз, удлиняется первая пара ног - появляются формы, близкие к var. cederstroemii. Осенью происходят, обратные изменения: укорачивается хвостовой придаток; уменьшаются коготки.
Литература
1. Бирштейн Я. А., Заренков Н. А. "Подкласс Жаброногие ракообразные (Branchiopoda)" / Жизнь животных. 2-е изд. Т. 2. Моллюски. Иглокожие. Погонофоры. Щетинкочелюстные. Иглокожие. Полухордовые. Хордовые. Членистоногие. Ракообразные. Москва, 1988. С.295-311
2. Semenchenko V. P. "Analysis of the dynamics of the numbers and stability of cladoceran populations in eutrophic lake" / Russian Journal of Ecology. Vol.27, No1. 1996 pp.41-46
3. Мануйлова Е. Ф. "Ветвистоусые рачки (Cladocera) фауны СССР". Москва-Ленинград, 1964. -328с.
4. Вежновец В. В. "Ракообразные (Cladocera, Copepoda) в водных экосистемах Беларуси: каталог. Определительные таблицы". Минск, 2005. -150с.