Eurytemora lacustris Poppe, 1887 (озерная эвритемора)
Отряд Calanoida. Семейство Temoridae.
Редко встречающийся в водной фауне Беларуси вид . Центр ареала - озера и дельты рек севера Европы и Зап. Сибири. Литве этот вид включен в Красную книгу, ранее был найден в 10 озерах, а в настоящее время встречается только в гиполимнионе 5 мезотрофных озер с максимальными глубинами от 38 до 60 м. Сокращение числа озер с E. lacustris в связи с изменением климата и эвтрофированием наблюдается и в соседней Латвии. В Польше вид зарегистрирован только в оз. Вигри. Показано, что вследствие эвтрофикации и повышения температуры в глубоких озерах вид исчезает. На территории Германии относится к редким и исчезающим видам.. В Беларуси отмечен только в двух мезотрофных озерах - Волчин (северо-запад Мядельского р-на Минской обл.) и Вечелье (Ушачский р-н Витебской обл.) бассейна р. Западная Двина. В оз. Вечелье Е. lacustris впервые обнаружен в июле 1976 г. По северо-западу Беларуси проходит южная граница ареала вида.
Длина тела взрослых особей в водоемах Беларуси от 1,5 до 1,7 мм, что несколько выше, чем указывается в справочниках. Туловище состоит из 3 отделов: головного (цефалона), грудного (торакса), слитых в головогрудь (цефалоторакс), и брюшного (абдомен). На конце абдомена две каудальные ветви с 5 оперенными щетинками. Головогрудь состоит из 5 сегментов, на которых размещены: пара длинных 25 -членистых антеннул, 5 пар ротовых конечностей и 5 пар плавательных ног. По внешнему виду похожа на лимнокалянуса, но отличается строением пятой пары плавательных ног и прикрепленным к абдомену самки яйцевым мешком.
Самка. Тело стройное, с длинным абдоменом. Обе ростральные нити умеренной длины, на концах закруглены. Цефалоторакс заметно суживающийся к заднему концу, с наибольшей шириной в переднем отделе; последний торакальный сегмент очень короткий, задние концы его мало выдаются, выростов не образуют. Соотношение длин цефалоторакса и абдомена равно 1,4-1,0. Генитальный сегмент абдомена заметно расширен в средней части и несет по краям несколько сенсорных волосков; наибольшая ширина сегмента превышает длину; генитальная пластинка сзади равномерно закруглена. 2-й абдоминальный сегмент лишь ненамного короче генитального, а анальный в 1-5 раза длиннее предыдущего. Каудальные ветви слабо расходятся на концах и опушены по внутреннему краю; длина их приблизительно равна длине 2 последних cerментов абдомена. На анальном сегменте и каудальных ветвях шипиков нет. Каудальные щетинки умеренной длины, обычно густо опушенные. Антеннулы почти достигают конца цефалоторакса. Околоротовые конечности и плавательные ноги 1-4-й пар обычного для рода строения. Базиподит 5-й пары ног широкий,суживающийся кзади, его наибольшая ширина больше длины, несет короткую голую щетинку. 1-й членик экзоподита с длинным внутренним выростом, направленным почти перпендикулярно продольной оси членика, заостренным на конце и вооруженным по заднему краю, в дистальной половине, мелкими шипиками; наружный край членика несет 2 одинаковых коротких и заостренных на вершине шипа. Терминальный членик примерно вдвое короче предыдущего, овальный, на конце несет 2 шипа, из которых наружный в 5 раз короче апикального, между этими шипами иногда можно видеть 1 очень маленький шипик. Длина самок 1,1-1,5 мм.
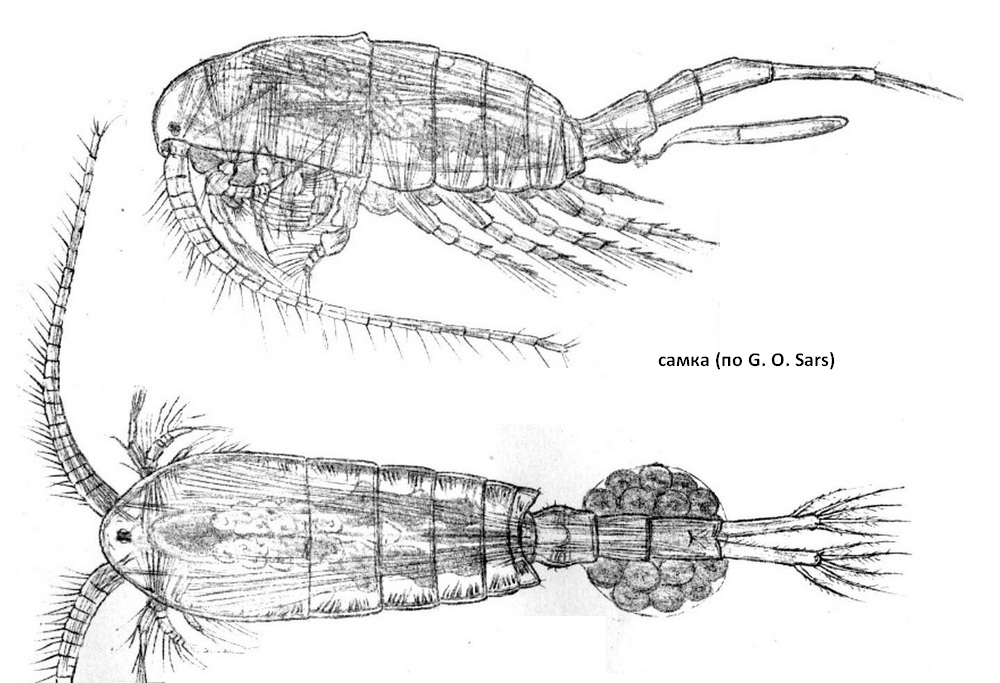
Самец. Тело тоньше и более стройное, чем у самки. Задние углы последнего торакального сегмента плавно округлены, достигают первой трети генитального сегмента абдомена. Генитальный сегмент с довольно толстым пальцевидным выростом слева. Каудальные ветви длинные (значительно длиннее и уже, чем у самки), параллельные, на концах не расходятся, внутренний их край опушен. Передняя и боковая поверхности анального сегмента и каудальных ветвей без вооружения. Эстетаски на левой антеннуле значительно длиннее, чем у самки. Правая антеннула геникулирующая; 8-12-й членики вооружены шипами, примерно одинаковыми по форме и длине. Ротовые части и 1-4-я пара плавательных ног устроены как у самки. Базиподит правой 5-й ноги удлиненный, заметно вздут совнутри и несет 2 тонкие щетинки - 1 на внутреннем и 1 на наружном крае. 1-й членик экзоподита длинный, с маленькой щетинкой в заднем отделе внутреннего края. Терминальный членик изогнут, несколько длиннее предыдущего, очень незначительно расширен при основании, несет 2 шипика на внутреннем крае и 1 тонкую щетинку на наружном крае. Базиподит левой ноги значительно короче, чем у правой, сильно вздут совнутри проксимально, иногда несет щетинку на передней поверхности, 1-й членик экзоподита примерно в 1,5 раза длиннее базиподита, вооружен 2 шипиками на передней поверхности. 2-й членик несколько длиннее предыдущего, сужен проксимально и сильно расширяется на конце, образуя двураздельную лопасть, большей частью вооруженную мелкими шипиками и волосками; на внутреннем и наружном краях и в основании наружной лопасти имеется по маленькому шипику. Длина самцов 1,0-1,4 мм.
Тело обоих полов почти прозрачно и совершенно бесцветно.
Редкий вид. Населяет глубокие мезотрофные озера с признаками олиготрофии. Живет в гиполимнионе озер, при низкой температуре воды. Встречается в планктоне круглый год, максимальное развитие популяций в летние месяцы. В июле найдены все возрастные стадии развития от науплиев до взрослых самок с яйцами. Средняя плотность в озерах Беларуси от 1380 до 2877 экз./м³. Максимальная плотность (до 12000 экз./м³) отмечена в оз. Вечелье в гиполимнионе. В 1950 г. средняя численность на глубоководной станции оз. Волчин составила 4400 экз./м³. Такой же показатель в этом озере в 1992 г. составил 1632, а в 2002 г. - 1380 экз./м³, что свидетельствует о снижении плотности популяции. Многолетние наблюдения на этих озерах показывают постепенное снижение прозрачности, основного показателя трофности озер. Особенно резко это проявляется в оз. Вечелье, где по данным 1976 г., летом этот показатель составлял 4 м. В 2005 г. прозрачность снизилась до 2, а в летний сезон 2011 г. составила только 1,2-1,5 м. Менее выражен процесс эвтрофирования в оз. Волчин, где снижение прозрачности за этот же период произошло только на 1 м - с 4,6 до 3,5, что находится в пределах межгодовых и сезонных колебаний этого показателя. На фоне снижения прозрачности воды наблюдается значительное падение численности популяции: за последние 30 лет почти в 8 раз. Так, средняя плотность E. lacustris в столбе воды, учитывая все возрастные стадии развития, в 1977 г. достигала 7,92 тыс. экз./м³, в 2002 была 2,9, а в 2005 только 1,01 тыс. экз./м³. Наблюдения 2011 г. показали дальнейшее снижение плотности до 0,22 тыс. экз./м³, что создает угрозу полного исчезновения этого вида из фауны озера. Известно, что процессы эвтрофирования озер создают неблагоприятные условия для обитания этого холодолюбивого стенотермного вида, проявляющиеся, прежде всего, в снижении уровня растворенного кислорода в гиполимнионе, где располагается основная часть популяции. Несмотря на падение прозрачности, в течение 2002-2011 гг. уровень растворенного кислорода в гиполимнионе снизился незначительно с 5,6 до 4,8 мг/л и оставался благоприятным для развития популяции.
В оз. Вечелье в летний период (июнь-июль 1976 г.) численность Е. lacustris была настолько малой, что он не встречался в количественных пробах. В августе численность рачка составила 0,09 тыс. экз/м³, биомасса 2,80 мг/м³. В планктоне присутствовали только взрослые особи. Самый высокий уровень плотности популяции отмечен в сентябре и декабре с максимальными значениями численности (7,92 тыс. экз./м³) и биомассы (46,10 мг/м³) в конце сентября. В сентябре 94,7% численности и 81,3% биомассы составляли науплиусы и только 5,3% численности и 8,7% биомассы приходилось на взрослых рачков и копеподитов. В феврале и марте численность популяций снизилась до минимума, причем в марте найдено всего три экземпляра копеподитной стадии - в двух качественных пробах. Зафиксировано также наличие науплиальных форм, взрослые рачки в марте отсутствовали.
Вертикальное распределение E. lacustris может определяться многими факторами (обилием пищи, содержанием растворенного кислорода, наличием рыб-планктофагов и др.). Многолетние наблюдения на озерах Беларуси показали, что популяция в летнее время постоянно располагается в гиполимнионе при температуре не выше 15°С, что свидетельствует о принадлежности озерной эуритеморы к холодолюбивой фауне. Вертикальное распределение науплиусов Е. lacustris отличается от распределения взрослых рачков и копеподитов в толще воды. На протяжении суток основная масса науплиусов пребывает в гиполимнионе. В зависимости от времени суток меняется их плотность относительно нижней границы термоклина (слой воды 1 м на глубине 11-12 м). Температурный скачок в термоклине 3,6°С (от 10,7°С у верхней границы до 7,1° С у нижней). В отличие от науплиусов вертикальные «миграции» копеподитных стадий и взрослых рачков происходят более активно. Суточные «миграции» науплиусов совершаются в основном в гиполимнионе, наибольшая плотность их меняется в диапазоне глубин от 12 до 16 м. «Миграции» взрослых рачков и копеподитов сходны. Эти две группы в больших количествах пересекают слой температурного скачка и достигают поверхности эпилимниона. Основная амплитуда суточных миграций составляет 8 м, т. е. в два раза больше, чем у науплиусов. Общей характерной особенностью всей популяции является то, что в первой половине светлого времени суток с 11 до 15 ч происходит передвижение популяции из верхних слоев в нижние с некоторым рассеянием плотности по всей толще воды. С наступлением темноты к 23 ч популяция снова уплотняется в вышележащих горизонтах и к 3 ч концентрируется у отмеченных для каждой группы верхних границ. В предрассветные часы наблюдается смещение наибольшей плотности популяционных групп от верхних границ к глубине. Взрослые особи и копеподиты в это время опускаются ниже термоклина и уплотняются на глубине 12 м, науплиусы опускаются по гиполимниону и уплотняются на глубине 16 м.
Размеры взрослых рачков в период с августа по февраль 1976 г. колебались в пределах 1,16-1,55 мм. При средних размерах 1,37 и 1,52 мм индивидуальный вес соответственно равен 0,030 и 0,032 мг. Размеры самцов немного меньше размеров самок. Индивидуальные веса особей копеподитных стадий при средних размерах 0,77, 0,97 и 1,17 мм соответственно 0,010, 0,017 и 0,025 мг. С февраля 2014 по февраль 2015 проведены новые исследования на оз. Вечелье. Размеры копеподитных стадий VI самцов колебались в пределах 1,092−1,534 мм (1,340±0,077), самок - 1,092−1,534 мм (1,308±0,069). Длина тела взрослых особей в озерах Беларуси сопоставима с размерами, указанным для озер России (1,0-1,4 мм) и некоторых стран Западной Европы - Польши (самки - 0,923-1,425 мм; самцы - 1,245-1,290 мм) и Швеции (самки 1,5-1,64 мм; самцы - 1,2 мм). Для большинства пресноводных каланоидных копепод характерны более крупные размеры самок. Но у E. lacustris средняя длина тела самцов больше на 0,021-0,056 мм за счет длины абдомена, что является отличительной особенностью вида.
За период исследований соотношение полов популяции в оз. Вечелье изменялось. Доля самцов во взрослой части популяции в июне составляла 92,6%. Примерно таким оставалось соотношение в июле с едва заметной тенденцией к увеличению доли самок. Резкий сдвиг наблюдался в августе, когда самцы лишь на 7,6% преобладали над самками. В середине сентября эта разница уменьшилась до 4%, а в конце сентября доля самцов понизилась до 46%. В декабре и феврале отмечалось дальнейшее увеличение разницы соотношения полов в пользу самцов. Полное исчезновение взрослых рачков обоих полов из популяции произошло в марте.
Доля яйценосных самок к середине сентября достигла 29,6% общей их численности. К концу сентября она резко снизилась и снова возросла к декабрю, затем постепенно снижалась до полного выпадения взрослых особей из популяции. Численность яйценосных самок в 2014-2015 гг. имела два достаточно выраженных минимума - в конце зимы и в конце лета. Максимумы численности фиксировались, соответственно, в конце весны (39,8 экз./м³), середине лета (48,7 экз./м³) и начале осени (28,7 экз./м³). Относительная плотность яйценосных самок также изменялась в течение года, принимая максимальные значения в конце зимы и середине лета (65,0-67,0%), а также в начале осени (79,3%), минимальное - в начале (22,9%) и конце лета (33,3%), а также поздней осенью (24,2%). В динамике изменения как абсолютной, так и относительной численности есть два наиболее выраженных пика, которые фактически совпадали по времени и приходились на середину лета и начало осени. Наличие в пробах фактически круглогодично самок с яйцами (а также сперматофорами) свидетельствует о растянутом процессе размножения. Кроме того, вероятно, что часть самок одной из генераций переходит в другую и участвует в размножении следующего поколения. Осенне-зимняя генерация более растянута во времени за счёт низкой температуры и неравномерного созревания особей.
Плодовитость отдельных самок в 1976 г. колебалась в пределах 3-43 яиц в кладке. Средняя плодовитость самок составляла в августе 27±1,2, в сентябре 34±0,9, в декабре 10±0,6, в феврале 6±0,8. В 2014-2015 гг. индивидуальная плодовитость E. lacustris в водоемах Беларуси при среднем значении - 13, изменялась от 8 до 48 яиц, была ниже величин из других точек ареала, для которых число яиц в яйцевом мешке указывается в пределах 32-56, а среднее значение составляет 44 яйца. Впервые для этого вида определены размеры яиц (средний диаметр составляет 100,53±1,73 мкм; мин. - 88, макс. - 112 мкм).
Как и все копеподы, E. lacustris имеет непрямое развитие и проходит 12 возрастов: шесть науплиальных (N), или личиночных, стадий и столько же копеподитных (C). Возрастной состав популяции значительно изменялся в сезоне. В зависимости от времени года и глубины наблюдалось значительное колебание соотношения науплиев и копеподитов в популяции. В целом же за годичный период процентное их соотношение в популяции приблизительно совпадало: личинки (52%) и копеподитные стадии развития (48%) вносят почти равноценный вклад в создание плотности популяции данного рачка. Так как данные 1976 г. несколько устарели, ниже приводятся только сведения по исследованиям 2014-2015 гг. в оз. Вечелье.
Науплии 1 (N I). Стабильно представлены в популяции с февраля по август. Наибольшая плотность и вклад в состав популяции наблюдались в июне-июле (192 экз./м³; максимальная доля в популяции - 13,4%). В августе произошло снижение доли и особенно численности стадии, вслед за чем в сентябре наблюдался новый пик (192 экз./м³ и 9,2% соответственно). До ноября шло постепенное падение, после чего абсолютная численность вплоть до февраля следующего года оставалась стабильно низкой, а доля в популяции к зимним месяцам несколько увеличилась и составила 5,7%. Таким образом, развитие этой стадии достигает максимума в июне-июле и затем в сентябре, наименее всего она представлена в ноябре-феврале. Размеры 0,120−0,136 мм (0,130±0,005).
Науплии 2 (N II). Представлены в популяции с февраля по ноябрь. Максимальное развитие этой стадии наблюдается в июне (633,8 экз./м³ и 27,9%) а также августе-сентябре (максимум численности наблюдался в сентябре - 547,8 экз./м³, доли в популяции - в августе (25,3%)). С сентября по ноябрь шло снижение доли и численности стадии, выраженное более сильно, чем у первых науплиусов. Размеры 0,136−0,184 мм (0,162±0,010).
Науплии 3 (N III). Постоянно представлены в популяции с февраля по ноябрь, создавая в этот период 3 чётко выраженных пика численности и доли, которые совпадают (май - 335,8 экз./м³ и 17,1%; июль - 360 экз./м³ и 26 %; октябрь - 509 экз./м³ и 23,1% соответственно). Вслед за спадом в ноябре численность увеличивалась очень медленно, в отличие от доли, которая резко возросла к январю, дав новый пик (16,1%, с тенденцией последующего снижения). Размеры 0,184−0,224 мм (0,199±0,013).
Науплии 4 (N IV). Вслед за зимним пиком развития (февраль) до конца мая шло постепенное снижение численности и доли стадии. Затем следовали 3 пика плотности и численности: менее выраженные в июне и второй половине июля, более выраженный - в сентябре-октябре (наиболее высокая численность и доля наблюдалась в начале октября - 575,9 экз./м³ и 26,2% соответственно, этот пик сопоставим с зимним). Размеры 0,232−0,280 мм (0,250±0,016).
Науплии 5 (N V). Наблюдается сходная динамика численности, как и у науплиев 4, с наиболее выраженными пиками зимой (февраль) и в начале осени (сентябрь - 407,3 экз./м³ и 15,9%). Размеры 0,280−0,344 мм (0,311±0,019).
Науплии 6 (NVI). По абсолютной численности наблюдались 2, по доле в зоопланктоне - 3 пика развития (конец мая - 108,1 экз./м³ и 5,5%; такой же по выраженности пик развития наблюдался в сентябре-октябре). С июня по первую половину августа науплии практически не фиксировались в озере. В январе-феврале вклад в состав популяции также был относительно высок для этой стадии (на уровне 3-6%), что не прослеживалось в динамике её численности. Размеры 0,352−0,408 мм (0,376±0,019).
Копеподиты 1 (С I). Динамика численности практически аналогична таковой у науплиев 6 (пик в мае - 158,6 экз./м³ и 7,8%; сентябрь - 139,2 экз./м³ и 5,4%). Со второй половины июня по первую половину августа копеподиты практически отсутствуют. В зимний период численность стадии находилась на довольно низком уровне. Размеры 0,442−0,546 мм (0,508±0,026).
Копеподиты 2 (СII). Также выражен поздневесенний пик развития (98,8 экз./м³ и 5,0%), но в отличие от 2 предыдущих стадий второй пик сильно растянут во времени и охватывает практически всю осень и зиму, хотя он и выражен довольно слабо (максимальная численность в ноябре - 145,2 экз./м³). Размеры 0,546−0,728 мм (0,648±0,038).
Копеподиты 3 (С III). Максимум первого всплеска развития приходится на конец весны и начало лета (189,7 экз./м³ и 8,3%), далее следует резкий спад, и всё лето стадия в водоёме не фиксируется. С приходом осени начинается второй всплеск развития, который достигает максимума в ноябре (916 экз./м³ и 24,6%) и заканчивается к началу января, в январе-феврале численность стадии минимальна. Размеры 0,702−0,910 мм (0,812±0,043).
Копеподиты 4 (С IV). Наблюдаемый первый рост численности растянут практически на весь весенне-летний период. В пределах его максимум численности и доли в популяции наблюдается к середине лета (200,9 экз./м³ и 17% соответственно). В первой половине осени стадия в озере практически не фиксировалась, а далее, как и у предыдущей стадии, следовал второй максимум в ноябре (994,5 экз./м³ и 25,4%) и снижение доли зимой. Размеры 0,832−1,118 мм (0,960±0,050).
Копеподиты 5 (С V). Первый всплеск развития также растянут на весь весенне-летний период, проявляя регулярные колебания. При этом максимум численности у самок приходится на начало лета (91%), самцов - на середину (85,8%). Далее наблюдается тенденция, как у предыдущей стадии, с той разницей, что второй пик выражен слабее (475,8 экз./м³ и 12,8% - самки, у самцов примерно те же величины численности). Размеры самцов 1,040−1,300 мм (1,164±0,052); самок 0,988−1,248 мм (1,099±0,082).
Копеподиты 6 (С VI). Животные этой стадии развития относятся к половозрелым особям. В годовой динамике численности этой стадии наблюдаются 2 пика: в начале лета (101,9 экз./м³ самки, 125,4 экз./м³ самцы) и в ноябре (189,7 экз./м³ у самцов, у самок он выражен лишь наполовину). Во второй половине лета и первой половине осени, а также во второй половине зимы стадия практически не фиксируется. Вклад же в состав популяции рачка у данной стадии максимален ближе ко второй половине лета и во второй половине зимы, когда относительная численность стадии колеблется в районе 5-7%. Размеры самцов 1,092−1,534 мм (1,340±0,077); самок 1,092−1,534 мм (1,308±0,069).
В годовой динамике зрелых особей популяции прослеживается чёткий волнообразный характер с 2 пиками и, соответственно, спадами их развития. Максимальная доля в популяции наблюдалась в июле 2014 и составила 13,8%. Зимний максимум 2015 года был немного меньше летнего (10,8%). Минимумы развития зафиксированы в конце февраля 2014 (1,3%) и в первой половине осени 2014 года (2,4-2,9%). Несовпадение значений численности в конце февраля 2014 и 2015 годов вызвано годовыми особенностями среды обитания и обусловлено температурой, которая могла сместить начало и сроки развития отдельных стадий. В целом, в течение года у E. lacustris наблюдаются две генерации.
Анализ возрастного состава копеподитной части популяции указывает на развитие двух генераций Е. lacustris. Это подтверждается наличием в пробах самок со сперматофорами с начала августа до конца сентября. Наибольшее число яйценосных самок и самок со сперматофорами отмечено в середине сентября, к концу этого месяца доля их значительно уменьшилась. Такой факт, очевидно, свидетельствует о летне-осенней генерации Е. lacustris, активный период которой проходит с августа по сентябрь. Наличие яйценосных самок и самок со сперматофорами в декабре и явное понижение их доли в феврале, вероятно, указывает на то, что в течение этого периода в оз. Вечелье протекает зимняя генерация Е. lacustris с наибольшей активностью в декабре - январе.
Судя по изменениям в возрастной структуре копеподитов и взрослых рачков, период развития копеподитов для зимней генерации около 1-1,5 месяца, а для копеподитов летне-осенней генерации он значительно короче. Вероятнее всего, июньские рачки являются поколением зимней генерации. Растянутый период размножения в зимнее время обусловливает одновременное присутствие в летней популяции копеподитов всех возрастов и неодновременное созревание, в результате летняя генерация оказывается растянутой на два месяца (август-сентябрь). Августовское поколение к декабрю достигает половозрелости и начинает зимнюю генерацию. Повышение доли копеподитов первой стадии в декабре подтверждает появление в популяции нового поколения летне-осенней генерации. Есть основание полагать, что декабрьские копеподиты первой стадии - молодь второй половины летне-осенней генерации, так как доля их через два месяца снизилась за счет переходов в более взрослые стадии. Взрослые рачки декабря, очевидно, поколение первой половины летне-осенней генерации.
На основании изменений в возрастном составе копеподитов первой стадии, можно предположить, что превращение науплиусов летне-осенней генерации, с момента выклева до копеподитов этой стадии, протекает около 1,5 месяцев, считая, что декабрьские копеподиты первой стадии - молодь, появившаяся в середине летне-осенней генерации. В конце марта в качественных пробах обнаружены только науплиусы Е. lacustris и несколько копеподитов третьей стадии, которые выросли к этому времени из науплиусов, появившихся в начале периода зимней генерации. Считая, что время перехода из одной копеподитной стадии в последующую занимает 1-1,5 месяца для копеподитов зимней генерации, можно считать, что примерный срок полного метаморфоза науплиусов зимней генерации до копеподитов первой стадии составляет 1,5-2 месяца, т. е. продолжительнее, чем у науплиусов летне-осенней генерации. Отсутствие взрослых рачков в мартовских пробах свидетельствует, очевидно, о том, что в феврале заканчивается цикл продолжительностью 6-7 месяцев, начавшийся в августе, а в августе-сентябре заканчивается цикл продолжительностью 8-9 месяцев, начавшийся в декабре. Можно говорить о дицикличности видовой популяции Е. lacustris в оз. Вечелье. По исследованиям 2014-2015 гг., дицикличность была подтверждена, но сделан несколько иной вывод - сроки развития обеих генераций составляют около 6 месяцев.
Наблюдения за озерной эвритеморой в рамках НСМОС начаты в 2008 году. С целью сравнения особенностей жизненного цикла было проведено углубленное изучение популяции в этом оз. Вечелье. Анализ возрастного состава в летние месяцы показал, что основу популяции составляют сложно определяемые науплиальные стадии развития. Более взрослые копеподитные стадии составляют около 20% популяции. Межгодовые изменения возрастного состава незначительны, что свидетельствует о стабильном состоянии популяции в этом водоеме. Прозрачность воды, измеренная по белому диску Секки, была 2,8 метра. Поверхностная температура воды составила 23,3°С, в эпилимнионе снижение температуры было незначительным, и на 5 м глубины составляла 21,5°С. Термоклин ярко выражен - с 5 до 6 м перепад температуры составил более 7°, что редко встречается в других стратифицированных озерах. Это связано с защитой озера от преобладающих ветров и его расположением в виде узкой полосы с юга на север. Резкое падение температуры наблюдалось до глубины 9 м. Таким образом, эту глубину можно считать концом термоклина. В дальнейшем, с глубиной, изменение температуры было плавным (клинолимнион), а начиная с глубины 18 м фактически оставалась неизменной - 5,1°С до самого дна. Таким образом, в этом водоеме для озерной эвритеморы, как холодноводного вида, только верхние 5 м являются непригодными для обитания, во всей остальной толще воды температурный фактор не является ограничивающим.
Численность зоопланктона в мезотрофном оз. Вечелье - около 100 тыс. экз./м³. Плотность популяции озерной эвритеморы составляет, в зависимости от года исследований, от 0,2 до 2,7 тыс. экз./м³. Например, по данным 2014-2015 гг. среднегодовая плотность Е. lacustris в столбе воды озера Вечелье составила 1939 экз./м³, минимальная - 654 экз./м³, максимальная - 3772 экз./м³, доля в зоопланктоне - 6,7%, среди копепод - 12,1%. Плотность в озере Волчин в 2–5 раз ниже, а доля в планктоне не превышала 2%. Из «мирных» каланоидных копепод со сходным типом питания и потенциальных конкурентов за пищевые ресурсы в планктоне оз. Вечелье присутствует только один вид – Eudiaptomus graciloides. В 2016 г. общая численность озерной эвритеморы в оз. Вечелье составила 1231 экз./м³, что находится в пределах межгодовых колебаний. До 80% популяции этого вида размещается в эпилимнионе. Пик плотности особей в 2377 экз./м³ наблюдался в слое воды 5-10 м и был обусловлен младшими стадиями развития. Взрослые особи концентрировались на глубинах 25-30 м. В 2017 г. при детальном учете всех стадий развития (науплиальных и копеподитных) численность в столбе воды составила 1209 экз./м³, взрослые животные составляли около 10% популяции и имели величину численности 118 экз./м³. Вертикальное дневное распределение E. lacustris в оз. Вечелье зависит от распределения температуры и возрастного состава. Младшие возрастные группы (науплии) держатся более высоких горизонтов, а взрослые, наоборот, обитают в глубоких слоях гиполимниона. В целом основная масса популяции озерной эвритеморы держится в мета- и гиполимнионе, а в прогреваемом приповерхностном слое 0-5 м встречаются только единичные особи, что свидетельствует о предпочтении низкой температуры. Максимум численности рачков в дневное время был приурочен к зоне температурного скачка, что объясняется, по-видимому, более богатыми кормовыми условиями для растущих молодых животных и расположен на глубинах 5-10 м. В 2017 г. средняя плотность взрослых животных была небольшой, поэтому, хотя их распределение значительно отличается от молоди, оно не влияет на общую картину распределения в целом. Пик численности старших копеподитных стадий развития был приурочен к слою с глубиной 25-30 м и температурой воды 5,1°С. Таким образом, по состоянию на 2017 год в оз. Вечелье сохраняется воспроизводимая и стабильная популяция эуритеморы озерной.
Пищевой комок E. lacustris содержит пеннатные виды диатомых, имеющих достаточно высокую плотность в гиполимнионе озер. Большинство водорослей повреждены - это свидетельствует об использовании «кремниевых коронок» режущего края мандибулы для их дробления и лучшего переваривания содержимого. Предположительно, эти водоросли принадлежат к виду Synedra pulchella, занимающему гиполимнион стратифицированных мезотрофных озер Беларуси.
К основным видам угроз для E. lacustris в этом водоеме, как и для всех животных, требовательных к качеству воды, относятся эвтрофирование и загрязнение, что ведет к снижению растворенного кислорода в толще воды, где она обитает. Одним из важных показателей эвтрофирования и загрязнения является прозрачность воды. Отмечено снижение этого показателя от 3-4 м (в середине прошлого века) до колебаний от 1,5 до 2,5 в настоящее время. На берегу озера нет значительных точечных источников загрязнения. Санаторий и льнозавод не функционируют. Рекреационное использование, несмотря на близость г. Ушачи, незначительна, озеро почти со всех сторон окружено лесом и хозяйственная нагрузка на водосборе тоже не может значительно влиять на экосистему водоема. Вероятно, могут сказываться последствия мелиорации, проведенной на водосборе, также направление течения поверхностных вод в южную часть озера. Учитывая высокие требования к качеству воды и содержанию растворенного кислорода в глубоких слоях воды для E. lacustris, при дальнейшем снижении прозрачности существует реальная угроза угнетения развития популяции.
Если не касаться Беларуси, то для вида указывается, что крупных водоемах он характеризуется многолетним жизненным циклом, максимум развития рачка приходится на холодную половину года; популяция держится в глубинных слоях воды.
Литература
1. Митрахович П. А., Ляхнович В. П. "Характеристика популяции Eurytemora Lacustris (Poppe) в озере Вечелье Белорусской ССР" / Вестник БГУ. Сер. 2. 1976. С.39-44.
2. Вежновец В. В. "Озерная эвритемора" / Красная книга Республики Беларусь. Животные: редкие и находящиеся под угрозой исчезновения виды диких животных. 2 изд. Минск, 2006. -318с.
3. Мониторинг животного мира / Национальная система мониторинга окружающей среды в Республике Беларусь: результаты наблюдений, 2016 год. С.299
4. Мониторинг животного мира / Национальная система мониторинга окружающей среды в Республике Беларусь: результаты наблюдений, 2017 год. С.326-327
5. Боруцкий Е. В., Степанова Л. А., Кос М. С. "Определитель Calanoida пресных вод СССР". Ленинград, 1991. -504с.
6. Вежновец В. В. "Ракообразные (Cladocera, Copepoda) в водных экосистемах Беларуси: каталог. Определительные таблицы". Минск, 2005. -150с.
7. Вежновец В. В., Зайдыков И. Ю., Наумова Е. Ю., Сысова Е. А. "Особенности биологии двух видов копепод (Crustacea, Copepoda, Calanoida) как возможные причины изменения их ареалов" / Российский Журнал Биологических Инвазий. № 2. 2012 С.16-29
8. Литвинова А. Г., Вежновец В. В. "Годовая динамика возрастного состава и цикл развития Eurytemora Lacustris (Poppe, 1887) в озере Вечелье (Беларусь)" / Вестник БарГУ. Выпуск 3, сентябрь, 2015. Серия «Биологические науки (общая биология). Сельскохозяйственные науки (агрономия)» С.34-43
9. Литвинова А. Г. "Биология и современное состояние популяций представителей рода Eurytemora (Copepoda, Calanoida) разного происхождения в водоемах Беларуси" / Автореферат диссертации на соискание уч. степени кандидата биол. наук. Минск, 2017. -24с.